 ,1,2,3,*, 朱敏1,2,3
,1,2,3,*, 朱敏1,2,3Subdivision and age of the Silurian fish-bearing Kuanti Formation in Qujing, Yunnan Province
CAI Jia-Chen1,2,3, ZHAO Wen-Jin,1,2,3,*, ZHU Min1,2,3通讯作者: *zhaowenjin@ivpp.ac.cn
收稿日期:2019-11-18网络出版日期:2020-10-20
| 基金资助: |
Corresponding authors: *zhaowenjin@ivpp.ac.cn
Received:2019-11-18Online:2020-10-20

摘要
云南曲靖地区的志留系出露较好,连续发育,化石丰富,是研究我国志留系理想的地区之一。自2007年在关底组中发现大量鱼类化石(2009年命名为潇湘脊椎动物群)以来,曲靖地区的关底组便成为国际上早期脊椎动物研究者密切关注的对象与焦点。然而从1914年关底组命名以来,其划分方案及地质时代归属等至今存有较大争议。结合近年来在潇湘水库周边地区详细的野外地质考察以及一条连续剖面(上铁路-东坡剖面)的实测资料,并在前人古生物学研究与地层学划分对比工作的基础上,对曲靖地区含丰富志留纪鱼化石的关底组重新进行了厘定与划分,并对其地质时代开展了分析讨论。根据主要岩性变化及化石特征,取消后期一些****从关底组下部划分出的岳家山组,并将其划分为4段,从下往上依次为关底组I段岳家山段、关底组II段崇家湾段、关底组III段彩莲段及关底组IV段东坡段。含潇湘脊椎动物群及丰富牙形类Ozarkodina crispa的关底组III-IV段的地层可归入到罗德洛统卢德福特阶,而含牙形类O. crispa的关底组II段上部及不含牙形类O. crispa的关底组I-II段中下部的地层目前暂归于罗德洛统高斯特阶为宜,但不排除其底部未见化石记录的关底组I段下部地层下延到温洛克世的可能。
关键词:
Abstract
The continuous Silurian strata are well developed and exposed with abundant fossils in Qujing Area of Yunnan Province, which makes Qujing as one of ideal areas in China for the research of Silurian stratigraphy and paleontology for a long time. The Kuanti Formation has become the focus of attention for early vertebrate researchers in the world, since tremendous amount of fossil fishes were found in the formation exposed in the surrounding areas of Qujing in 2007, which eventually led to the discovery and establishment of the Xiaoxiang Vertebrate Fauna in 2009. However, the stratigraphic subdivision, correlation and the geological age of the Kuanti Formation in Qujing still remain contentious, although many biostratigraphic and lithostratigraphic attempts have been made since the formation was named in 1914. Based on the detailed field geological investigation in recent years in the surrounding areas of Xiaoxiang Reservoir and the measured data of one continuous section (Shangtielu-Dongpo Section), together with many previous paleontological and stratigraphic works, this paper redefines the Kuanti Formation containing abundant Silurian fishes in Qujing Area and further discusses its geological age. According to the main lithological changes and paleontological characteristics, the Yuejiashan Formation which was separated by some researchers from the lower part of the Kuanti Formation is abandoned here. In the paper, the redefined Kuanti Formation can be subdivided into four members in ascending order. Member I (Yuejiashan Member) is characterised by yellow-green and gray-green shales intercalated with thin-bedded fine sandstones or siltstones and several thin-bedded fine-grained conglomerates bearing fragments of fossil fishes in its lower and middle parts. Member II (Chongjiawan Member) is represented by gray-green and purple-red shales, intercalated with light grey middle- to thin-bedded or lenticular limestones or bioclastic limestones containing many brachiopod fossils. Member III (Cailian Member) is dominated by purple-red and gray-green silty and calcareous mudstones or marls intercalated with minor purple-red or yellow-green shales or siltstones, containing brachiopods, fossil fishes and stout tubular trace fossils. A set of middle-thick-bedded fine sandstone with small thickness is usually developed in the bottom of the member, which becomes the obvious marker of the boundary between Member III and Member II. Member IV (Dongpo Member) is composed of gray-green and yellow-green mudstones and shales intercalated with thin-bedded or lenticular argillaceous limestones and marlites. Abundant fossil fishes of the Xiaoxiang Vertebrate Fauna were found in the argillaceous limestones in the lower part, and coral fossils in the upper part. Mainly based on the records of fossil fishes, conodonts and other paleontological data, the age of Member III to Member IV of the Kuanti Formation, containing the main fish-bearing strata of the Xiaoxiang Vertebrate Fauna and conodont Ozarkodina crispa, should be assigned to the Ludfordian Stage of the Ludlow, and Member I to Member II can be referred to the Gorstian Stage of the Ludlow, Silurian. Based on the current stratigraphic data, the possibility of its bottom extending down to Wenlock is not excluded.
Keywords:
PDF (5890KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
蔡家琛, 赵文金, 朱敏. 云南曲靖志留纪含鱼地层关底组的划分与时代. 古脊椎动物学报[J], 2020, 58(4): 249-266 DOI:10.19615/j.cnki.1000-3118.200513
CAI Jia-Chen, ZHAO Wen-Jin, ZHU Min.
云南曲靖地区的志留系特别发育,且出露较好、化石丰富,是我国研究志留系较为理想的地区之一(方润森等,1985)。1914年,丁文江最早开展了曲靖地区志留系的野外地质调查工作,其研究成果发表于1937年(Ting and Wang, 1937)。此后70年来众多研究者对本区志留纪地层及其中所含的腕足类、珊瑚、三叶虫、介形虫、牙形类、早期脊椎动物等做了大量的研究(Wang, 1944, 1947; Sun and Wang, 1946; Yin, 1949; 伍鸿基,1977; 戎嘉余、杨学长,1978, 1981; 葛治洲等,1979; 王钰等,1980; 王成源,1980, 1981, 1998, 2001; 陈均远等,1981; 方润森等,1985; 潘江,1986; Pan and Dineley, 1988; 丁春鸣,1988; Walliser and Wang, 1989; 王念忠、董致中,1989; 王尚启等,1992; 曹仁关,1994; Geng et al., 1997; Zhu and Schutlze, 1997; 何心一、陈建强,2004), 取得了一系列的科研成果和进展,为本区志留系各岩石地层单位地质时代的确定以及洲际间地层对比提供了比较可靠的古生物学依据(戎嘉余、陈旭,1990)。然而曲靖地区的关底组自1937年正式命名以来,其划分方案及地质时代归属等地层学问题至今仍存有较大争议。
2007年以来,我们主要围绕志留纪鱼类化石的发现与发掘,在曲靖地区尤其是志留系发育较好的潇湘水库周边开展了大量的地质考察工作,不仅在志留纪鱼类化石的发现与研究中取得了系列重要进展,而且在志留纪含鱼化石海相地层特别是关底组的时空分布、划分与对比等方面取得了一些新认知。前人曾报道在曲靖地区关底组中发现了盾皮鱼类大化石(潘江,1986; Pan and Dineley, 1988)以及少量的鱼类微体化石(王念忠、董致中,1989), 不过这些盾皮鱼类大化石迄今仍未被描述。在近10余年的野外发掘工作中,我们在关底组中发现了数个鱼类大化石层位,其中除盾皮鱼类以外,还有一些盔甲鱼类、早期硬骨鱼类等材料。这些潇湘脊椎动物群鱼化石的发现,不仅丰富了我国古生代中期特别是志留纪的鱼群面貌,为深入探讨鱼类的系统演化如有颌类的早期演化和硬骨鱼类的起源等提供了重要的化石实证(Zhu et al., 2009, 2012, 2013, 2016; Choo et al., 2014, 2017), 还为含丰富鱼化石的关底组划分与对比以及时代归属等地层学问题提供了古鱼类学方面的新证据。
本文结合近年来在潇湘水库周边地区详细的野外地质考察以及一条连续剖面(上铁路-东坡剖面)的实测资料,并在前人古生物学研究与地层学划分对比工作的基础上,主要依据岩性变化及古生物化石特征,对曲靖地区含丰富志留纪鱼化石的关底组重新进行了厘定与划分,并对其地质时代开展了深入讨论。
1 剖面描述
位于潇湘水库西侧的上铁路-东坡剖面(图1A)志留系出露完好,古生物化石特别是腕足类、珊瑚及鱼类化石相当丰富,我们曾于2007年踏勘了此剖面,2017和2019年分两次实测了该剖面(图1B), 剖面详细描述如下。图1
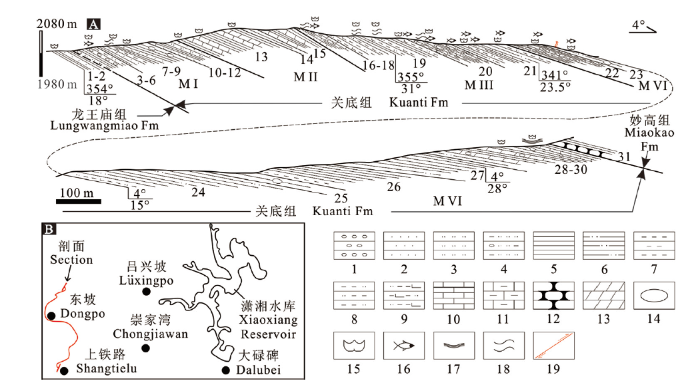 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1曲靖周边上铁路-东坡志留系关底组实测剖面及剖面位置
Legends: 1. 砾岩 conglomerate; 2. 细砂岩 fine sandstone; 3. 粉砂岩 siltstone; 4. 含砾泥质粉砂岩conglomerate-bearing argillaceous siltstone; 5. 页岩 shale; 6. 粉砂质页岩 silty shale; 7. 泥岩 mudstone;8. 粉砂质泥岩 silty mudstone; 9. 钙质粉砂质泥岩 calcareous silty mudstone; 10. 薄层灰岩 thin-bedded limestone; 11. 泥质灰岩 argillaceous limestone; 12. 瘤状灰岩 nodular limestone; 13. 泥灰岩 marlite;14. 透镜体 lens; 15. 无脊椎动物化石 invertebrate fossils; 16. 鱼化石 fossil fishes; 17. 叠层石 stromatolite;18. 遗迹化石 trace fossils; 19. 逆断层 reverse fault
Fig. 1The measured Shangtielu-Dongpo Section (A) of the Silurian Kuanti Formation around Qujing and its locality (B)
妙高组:
31. 浅灰色瘤状灰岩与黄绿色页岩互层,含腕足类、双壳类、腹足类、珊瑚等。未见顶


关底组(881.92 m):
关底组IV段(356.04 m)
30. 黄绿色页岩夹灰黄色灰岩、灰黑色泥灰岩薄层或透镜体,顶部灰岩或泥灰岩中含叠层石、珊瑚等,底部灰岩、泥灰岩薄层或透镜体中含大量生物碎屑 84.12 m
29. 浅灰色、褐黄色泥灰岩夹浅灰色灰岩薄层或透镜体,顶部夹少量灰黑色钙质页岩,底部夹少量黄绿色页岩 38.71 m
28. 土黄色粉砂质页岩夹薄层状灰岩、泥灰岩 9.71 m
27. 土黄色页岩夹灰白色页岩 54.08 m
26. 黄绿色页岩,底部夹少量同色钙质泥岩 46.23 m
25. 黄绿色页岩夹灰岩或泥灰岩透镜体 7.40 m
24. 黄绿色粉砂质页岩,底部夹极少量肉红色粉砂质页岩 63.94 m
23. 黄绿色粉砂质页岩与青灰色粉砂质泥质灰岩互层,夹少量黄绿色页岩,顶部夹中薄层黄绿色粉砂质钙质泥岩。本层上部及下部泥质灰岩中均含少量Lingula sp.和丰富鱼化石,是潇湘脊椎动物群产出的主要层位 11.87 m
22. 上部为黄绿色粉砂质钙质泥岩夹黄绿色粉砂质页岩,局部夹灰岩团块、薄层灰岩条带或灰岩透镜体,并含少量Lingula sp.和鱼化石;下部为黄绿色粉砂质页岩或粉砂质钙质页岩夹同色粉砂质钙质泥岩及灰岩透镜体或薄层生物碎屑灰岩,灰岩中含丰富腕足类化石 39.98 m
关底组III段(203.95 m)
21. 黄绿色粉砂质页岩夹紫红色及杂色粉砂质钙质泥岩,局部夹浅灰色瘤状灰岩薄层或透镜体。黄绿色、紫红色粉砂质钙质泥岩含Lingula sp., 杂色粉砂质钙质泥岩距本层底18.49 m处含盾皮鱼骨片、距本层底50.82 m处含鱼类鳞片化石 53.86 m
20. 黄绿色或灰绿色粉砂质钙质泥岩与黄绿色粉砂质页岩互层,夹杂色(紫红色与黄绿色为主)粉砂质钙质泥岩、灰岩透镜体或瘤状灰岩薄层。黄绿色粉砂质钙质泥岩中含大虫管和腹足类化石,杂色粉砂质钙质泥岩中含Lingula sp.和鱼化石,4个主要含鱼化石的层位距本层底分别为5.13, 21.33, 24.91和35.07 m 36.38 m
19. 紫红色、灰紫色粉砂质钙质泥岩夹紫红色粉砂质页岩、黄绿色粉砂质钙质页岩或粉砂质钙质泥岩。紫红色粉砂质钙质泥岩中含小虫管及Lingula sp., 距本层底50.48 m处黄绿色粉砂质钙质泥岩中含少量盾皮鱼类化石 80.82 m
18. 土黄色粉砂质钙质泥岩与灰黄色泥灰岩互层,普遍含大虫管,泥灰岩中含Lingula sp.和硬骨鱼类骨片 7.22 m
17. 黄绿色粉砂质钙质泥岩夹紫红色、黄绿色泥质粉砂岩与粉砂质页岩。上部为黄绿色粉砂质钙质页岩夹粉砂岩,顶部为浅黄色粉砂岩;中部为黄绿色粉砂质钙质泥岩,含少量Lingula sp.和鱼化石;下部为紫红色泥质粉砂岩夹粉砂质钙质页岩 16.45 m
16. 浅黄色中厚层状细砂岩夹黄绿色粉砂质页岩、紫红色页岩。顶部为中厚层状浅黄色细砂岩夹黄绿色粉砂质页岩,页岩中具大虫管;中部为紫红色页岩夹中薄层状浅黄色细砂岩;下部为厚层状浅黄色细砂岩 9.22 m
关底组II段(178.11 m)
15. 黄绿色、紫红色页岩互层,夹灰岩、泥灰岩薄层,顶部为浅黄色页岩 56. 31 m
14. 紫红色页岩与黄绿色页岩互层 21.22 m
13. 紫红色页岩与黄绿色页岩互层,夹灰岩、泥灰岩薄层或透镜体,含腕足类、海百合茎及珊瑚等化石 100.58 m
关底组I段(143.82 m)
12. 黄绿色页岩、薄层状黄绿色粉砂岩和土黄色细砂岩组成的韵律存在多个沉积旋回,向上粉砂岩、细砂岩减少,顶部夹褐色灰岩薄层,含腕足类小莱采贝(Retziella uniplicata)等化石 12.32 m
11. 黄绿色粉砂质页岩,中上部因覆盖未见原生露头 16.27 m
10. 黄绿色钙质粉砂质页岩、泥质粉砂岩与薄层状土黄色细砂岩互层,中部细砂岩层面上波痕发育 11.24 m
9. 黄绿色、灰青色页岩,中上部夹薄层状土黄色细砂岩,顶部砂岩层减少,夹钙质粉砂岩及细砂岩透镜体,含大量腕足类Retziella uniplicata等化石 39.66 m
8. 黄绿色页岩夹薄层状黄绿色粉砂质泥岩、黄褐色细砂岩,底部为一薄层泥岩,普遍含小虫管和大量腕足类Retziella uniplicata等化石 15.11 m
7. 灰绿色、黄绿色页岩与薄层状细砂岩互层,底部夹一薄层细砾岩,顶部夹一层黄绿色细砂岩 15.57 m
6. 黄绿色页岩与灰绿色页岩互层,含腕足类Lingula sp. 13.68 m
5. 灰绿色页岩夹细砾岩薄层,细砾岩中含少量鱼类微体化石,普遍含腕足类Retziella uniplicata等化石 7.47 m
4. 黄绿色页岩夹土黄色泥岩、粉砂质泥岩、泥质粉砂岩,含大量腕足类化石,底部含少量双壳类化石 4.83 m
3. 土黄色、黄绿色含细砾粉砂质泥岩夹灰白色、紫红色含砾泥质粉砂岩及土黄色细砾岩。上部为土黄色泥岩夹含细砾粉砂质泥岩或细砾岩;中部为中薄层状紫红色或黄绿色粉砂质泥岩,夹薄层状黄绿色含细砾粉砂质泥岩;下部为灰白色中层状含细砾泥质粉砂岩 7.67 m
龙王庙组:
2. 土黄色页岩夹薄层状灰白色细砂岩,底部为薄层状浅黄色细砂岩 17.62 m
1. 土黄色页岩,含三叶虫化石 4.42 m
2 关底组的厘定
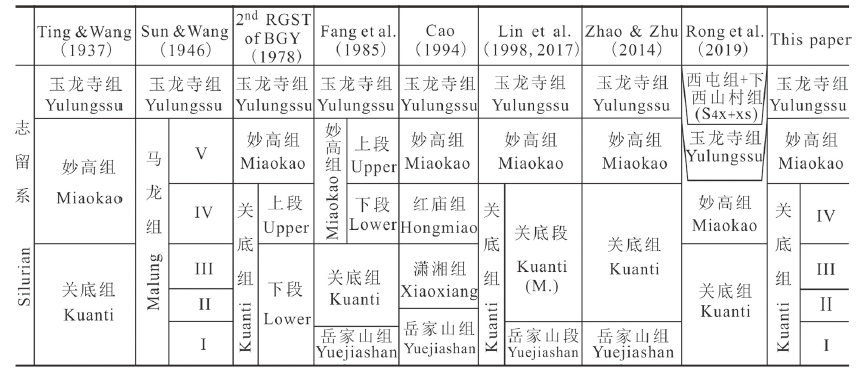
1914年,丁文江最早对曲靖地区发育的志留系开展了详细的野外地质调查工作,将之统称为马龙系,并划分出关底组、妙高组和玉龙寺组三个岩石地层单位(Ting and Wang, 1937; 表1), 其中的关底组命名于滇东马龙县关底村至寻甸县易隆之间,参考剖面位于曲靖城西南的岳家山,以紫红、黄绿或灰绿色泥质粉砂岩、泥岩为主,夹细砂岩、泥灰岩,即下砂岩层(Lower Sandstone), 与产丰富腕足类的中灰岩层(Middle Limestone)即妙高组整合接触。Sun and Wang (1946)曾推断关底组与妙高组可与英国的Wenlock地层相对比,且依据其与上覆的玉龙寺组在岩相的横向变化上存在明显差异的特点,认为丁、王二氏所划分出的关底组与妙高组两个岩石地层单位似乎有些随意,遂依据1942年实测的岳家山-妙高山剖面将关底组与妙高组合并成一个组,并命名为马龙组。现在看来,关底组与妙高组的岩相在横向及纵向变化上亦表现得非常明显,丁、王二氏(1937)较孙、王二氏(1946)的划分方案更为可取。Table 1
表1
表1云南曲靖地区志留系划分对比
Table 1
 |
新窗口打开|下载CSV
1960年代起,众多国内****先后对曲靖周边出露的志留系开展了大量的古生物地层学方面的工作,关底组的定义与划分也几经变化、争议不断(表1)。1964年云贵石油勘探处102队将丁、王二氏定义的关底组下部非红层的岩性段单独划分出来并命名为岳家大山群,该群随后常被称为岳家山组(林宝玉等,1984; 方润森等,1985; 云南省地质矿产局,1990; 曹仁关,1994), 而其上部仍称为关底组(方润森等,1985), 这实际上缩小了关底组下限的最初定义范围。1978年,云南省地质局第二区域地质测量大队在完成1:20万区域地质调查报告“曲靖幅”时,对曲靖潇湘水库地区的志留系进行了实测,虽仍分为关底组、妙高组和玉龙寺组,但这些地层单位的涵义已发生一些变化(表1), 以瘤状灰岩的大量出现作为关底组与上覆妙高组的区分界线,实际上扩大了关底组上限的最初定义范围,此种划分方案得到了一些后期研究者的支持(葛治洲等,1979; 丁春鸣,1988; 云南省地质矿产局,1990; 林宝玉等,1998, 2017; 表1)。曹仁关(1994)曾根据岩性、化石和岩相等特征将曲靖地区的志留系重新进行了划分,除玉龙寺组采纳了原始定义外,妙高组及关底组均二分,分别包括妙高组、红庙组与潇湘组、岳家山组(表1), 此方案给曲靖周边地区本就争议不断的志留系划分进一步造成了困扰。黄冰等(2011)与周航行、黄冰(2018)在研究黔西赫章志留纪晚期小莱采贝动物群(Retziella Fauna)及曲靖地区志留纪无洞贝Atrypoidea foxi Jones居群时,认为曲靖地区妙高组之下的地层从下到上的岩性特征相似、地质时代相同,且腕足动物等无脊椎动物化石组合面貌无本质差异,建议取消一些****将其下部单独划分出岳家山组,并指出仅用关底组称之的意见应更为合理,王成源(2013)及戎嘉余等(2019)均持此观点(表1)。赵文金、朱敏(2014)则采纳了林宝玉等(1984)将非红层的关底组下部单独划分出来并称之为岳家山组[林宝玉等(1998)又改称岳家山段]的划分意见(表1)。
综上所述,曲靖地区的志留系关底组自其命名以来,众多国内****对其涵义的理解及划分的具体方案并未获得一致的意见,争议的焦点表现在以下两个方面:一是其下界的定义,是否将其下部单独划分出岳家山组并将关底组的下界定义在两组的界线之间?二是其上界的定义,是以瘤状灰岩的大量出现作为关底组与上覆妙高组的区分界线,还是以中薄层灰岩夹层或透镜状灰岩的出现作为上覆地层妙高组的开始?
《中国地层指南及中国地层指南说明书》(2001)指出,组是岩石地层划分的基本单位,是野外宏观岩类或岩类组合相同、结构类似、颜色相近、呈现整体岩性和变质程度特征一致、空间上有一定延展性的地层体;组主要以地层的岩石特征和岩石类别作为划分依据,通常不考虑时间、成因、气候、环境或事件等因素;其岩石组合可由一种岩石构成,或者以一种主要的岩石为主、夹有重复出现的夹层,或者由两三种岩石互层构成,还可能以很复杂的岩石组分或独特的结构所构成并与其他比较单纯的组相区别;组的界线应为清楚、稳定的特殊岩性变化面或特殊结构构造标志层为界线划分。据此,在曲靖地区丁、王二氏(1937)、“曲靖幅” 1:20万区域地质调查报告(1978)、丁春鸣(1988)等志留系的三分方案至今仍有可取之处,将以海相碎屑岩为主并伏于以瘤状灰岩为主的妙高组之下的地层定义为关底组不仅符合《中国地层指南及中国地层指南说明书》(2001)的要求,而且在沿袭关底组历史沿革的基础上可以很好地解决目前有关其涵义的纷争。
结合近年在曲靖潇湘水库周边开展的大量野外地质工作,并在众多前人相关研究的基础上,笔者重新厘定了志留系关底组的涵义:以细碎屑岩为主的一套滨浅海相沉积,主要岩性以黄绿色、灰绿色、紫红色页岩、粉砂质页岩、粉砂质钙质泥岩、泥质粉砂岩为主,夹泥灰岩、灰岩及细砂岩薄层,富含腕足类、珊瑚、双壳类、介形虫与鱼类化石。本组在滇东地区广泛出露,厚度变化较大,通常在230~930 m之间,其与下伏下寒武统龙王庙组或中寒武统双龙潭组呈平行不整合接触,与上覆妙高组连续沉积并以浅灰色瘤状灰岩的出现作为妙高组的开始。
3 关底组的划分
重新厘定的关底组总体上代表了一套以滨浅海相红色细碎屑岩为主的沉积,其上、下部分的岩性组合特征并不完全相同、存有一定的差别,笔者主要依据岩性变化特点进一步将关底组划分成4段,由下往上依次如下。3.1 关底组I段——岳家山段
本段岩性主要以灰绿色、黄绿色页岩夹土黄色薄层状细砂岩为主(图2A), 其下部为黄绿色或土黄色(可能为风化色)页岩夹灰绿色细砾岩或含细砾粉砂岩、粉砂质泥岩薄层,靠近其顶部的砾石层中常含少量鱼类微体化石;中部为黄绿色与灰绿色页岩互层,夹土黄色细砂岩,普遍含大量腕足类化石,在上铁路-东坡剖面东侧崇家湾一带出露的细砂岩中含少量盾皮鱼类化石;上部为灰绿色页岩、黄绿色粉砂岩及细砂岩互层,偶夹紫红色页岩,普遍含腕足类化石,细砂岩层面常发育波痕。紫红色页岩的大量出现作为其上关底组II段的开始。本段在上铁路-东坡剖面上出露厚度为143.82 m, 与《云南省区域地质志》(1990)所描述的潇湘水库剖面中的岳家山组(厚129.3 m)层位相当,因此本文中将关底组I段称为岳家山段。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2曲靖周边上铁路-东坡剖面的志留系关底组地层
A. 关底组I段(Member I); B. 关底组II段(Member II); C. 关底组III段(Member III); D. 关底组IV段(Member IV);E. 关底组III段下部粗大虫管遗迹化石(the stout tubular trace fossils in the lower part of Member III);F. 关底组Ⅲ段中部盾皮鱼类化石(placoderms in the middle part of Member III)
Fig. 2The Silurian Kuanti Formation exposed in the Shangtielu-Dongpo Section around Qujing, Yunnan
3.2 关底组II段——崇家湾段
本段岩性以紫红色和黄绿色页岩互层为主,夹浅灰黑色灰岩、泥灰岩、钙质泥岩薄层或透镜体(图2B), 含丰富腕足类以及少量海百合茎、珊瑚等化石。本段以紫红色页岩的大量出现以及常夹灰岩、泥灰岩薄层等岩性特征可与下伏岳家山段相区分。本段在崇家湾-上铁路一带以及潇湘水库大坝附近均有出露,在崇家湾村北最为发育,遂将此段称为崇家湾段,在上铁路-东坡剖面上本段地层出露厚度为178.11 m。3.3 关底组III段——彩莲段
本段岩性以紫红色和黄绿色粉砂质钙质泥岩为主,夹同色页岩以及少量浅黄色细砂岩与浅灰黑色泥灰岩、瘤状灰岩薄层(图2C), 含大虫管以及腕足类、腹足类与鱼化石。本段底部为中厚层浅黄色细砂岩夹紫红色、黄绿色粉砂岩、页岩薄层,由下向上砂岩层减薄、页岩层增多,含大虫管、Lingula和少量鱼化石;下部为黄绿色粉砂质泥灰岩夹紫红色钙质泥岩、页岩,泥灰岩层中具大虫管(图2E)并含Lingula及鱼骨片化石;中部为紫红色钙质泥岩或泥灰岩夹紫红色、黄绿色页岩、黄绿色钙质泥岩,向上红层减少,以黄绿色或灰绿色为主,并夹少量灰岩透镜体或瘤状灰岩薄层,含大虫管、腕足类、腹足类和鱼化石(图2F); 上部为黄绿色、浅黄绿色页岩为主,夹紫红色、灰紫色页岩及黄绿色、紫红色钙质泥岩、泥灰岩和灰岩透镜体,含腕足类及鱼化石。本段在曲靖市区西南潇湘水库周边地区特别是彩莲村南非常发育且化石丰富,因此在本文中将此段称为彩莲段。本段在上铁路-东坡剖面上的地层出露厚度为203.95 m, 其底部的中层状细砂岩可以作为与II段——崇家湾段相区分的良好标志层,而红层的大量减少及至消失则可看作本段的结束。3.4 关底组IV段——东坡段
本段岩性主要以黄绿色粉砂质页岩、钙质泥岩、粉砂岩为主,夹浅灰色、灰黑色灰岩条带、灰岩薄层或透镜体(图2D), 含腕足类、珊瑚以及鱼化石。下部为黄绿色粉砂质页岩夹黄绿色粉砂质泥岩、灰岩薄层及泥质灰岩透镜体,含腕足化石和鱼化石;中部为黄绿色页岩夹灰岩、泥灰岩薄层或透镜体;顶部为黄绿色页岩夹泥灰岩、灰岩透镜体或薄层,含叠层石、珊瑚化石。本段在曲靖市区西南潇湘水库周边地区特别是东坡村一带非常发育且化石丰富,因此在本文中将此段称为东坡段。本段地层与1:20万区域地质调查报告“曲靖幅”中所定义的关底组上段、方润森等(1985)所定义的妙高组下段一致。在上铁路-东坡剖面上,本段地层出露厚度为356.04 m, 其下部泥灰岩透镜体是潇湘脊椎动物群的主要产出层位。本段与下伏III段——彩莲段可以红色海相地层的消失以及灰岩薄层、泥质灰岩透镜体的出现作为很好的区分标志,而其顶则可以大量瘤状灰岩的出现作为本段的结束、上覆妙高组的开始。4 关底组的鱼化石及其产出层位
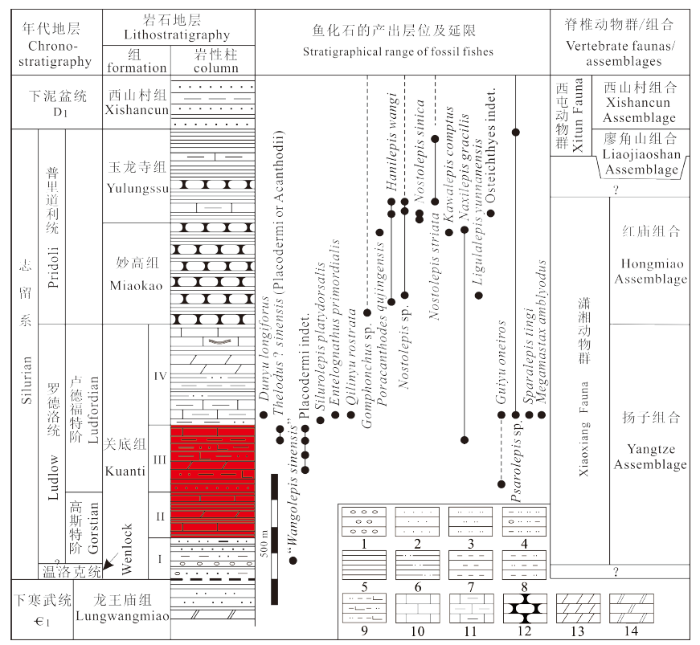
曲靖周边地区的志留系关底组十分发育、古生物化石丰富,除含丰富的腕足类、珊瑚、双壳类、介形虫、牙形类等无脊椎动物化石以外,脊椎动物化石也相当丰富,以盔甲鱼类、盾皮鱼类、棘鱼类以及硬骨鱼类为代表的潇湘脊椎动物群的发现与建立使得曲靖地区的关底组成为全球古脊椎动物学家瞩目的焦点。曲靖周边地区已不仅在早泥盆世时期是鱼类的天堂,而且在志留纪期间也成为鱼类的故乡。曲靖周边地区关底组鱼化石的发现可追溯到1970年代。云南省地质局第二区域地质测量大队在完成1:20万区域地质调查报告“曲靖幅”时,于1976年首先在大碌碑至吕兴坡公路西侧关底组底部薄层细砾岩(位于本文中关底组I段)中发现鱼化石碎片,随后分别在寻甸县麦厂街东南关底组内距底约30 m的黄绿色泥质粉砂岩(位于本文中关底组I段)、潇湘水库大坝附近关底组中部(相当于本文中关底组III段)紫红色钙泥质粉砂岩中发现并采获一些鱼化石(云南省地质局第二区域地质测量大队,1978), 但可惜这批鱼化石材料至今均未描述发表。潘江(1986)在“中国志留纪脊椎动物群的初步研究”一文中提及了采于寻甸县麦厂街附近的盾皮鱼类化石,并将其命名为“中华王氏鱼”(“Wangolepis sinensis”) (图3), 但迄今仍一直以裸名的方式在文献中被提及,并未被详细描述(赵文金、朱敏,2014)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3滇东志留纪鱼化石的地层分布
Legends: 1-13. 见
Fig. 3Stratigraphical distribution of Silurian fossil fishes from East Yunnan
王念忠、董致中(1989)最早描述并报道了曲靖周边志留系中的鱼类微体化石,包括采自关底组III段——彩莲段中的Thelodus sinensis和Naxilepis gracilis等(图3)。其中的Thelodus sinensis经重新观察或者是盾皮鱼类的外骨骼瘤点残片,或者是棘鱼类的鳍刺,而不是花鳞鱼类(M?rss et al., 2007)。随后,Wang (1993)曾提及了采自关底组上部(位于本文中关底组IV段下部)的一个躯甲保存较好的盾皮鱼类,当时将之称为阔背志留鱼(Silurolepis platydorsalis) (图3), 但直到2010年相关的形态学描述才完成(Zhang et al., 2010), 并在最近完成了更深入的研究工作(Zhu et al., 2019)。
2007年,中国科学院古脊椎动物与古人类研究所早期脊椎动物研究团队开始在曲靖周边千余米厚的志留纪地层中开展详尽的地质考察与化石发掘,在关底组上部——IV段东坡段下部的青灰色泥灰岩中发现并采获了大量的鱼类化石材料。在这批早期脊椎动物化石中,既有大型的盾皮鱼类,又有以胴甲鱼类为代表的小型盾皮鱼类,此外还包括大量原始的硬骨鱼类、盔甲鱼类等早期鱼类类群,2009年随着梦幻鬼鱼(Guiyu oneiros)的研究报道将之称为潇湘脊椎动物群(Zhu et al., 2009), 随后的研究又将其细分为以硬骨鱼类、盾皮鱼类、盔甲鱼类等大化石为代表的扬子鱼类组合和以棘鱼类、硬骨鱼类微体化石为主的红庙鱼类组合(Zhao and Zhu, 2010, 2014, 2015; 图3)。迄今,关底组潇湘脊椎动物群中已报道的鱼类大化石主要包括:硬骨鱼类梦幻鬼鱼(Guiyu oneiros Zhu et al., 2009)、钝齿宏颌鱼(Megamastax amblyodus Choo et al., 2014)、丁氏甲鳞鱼(Sparalepis tingi Choo et al., 2017), 盾皮鱼类初始全颌鱼(Entelognathus primordialis Zhu et al., 2013)、长吻麒麟鱼(Qilinyu rostrata Zhu et al., 2016)、阔背志留鱼(Silurolepis platydorsalis Zhang et al., 2010)、“中华王氏鱼”(“Wangolepis sinensis” 潘江,1986), 以及盔甲鱼类长孔盾鱼(Dunyu longiforus Zhu et al., 2012)。除“中华王氏鱼”采自关底组I段中部外,其他重要鱼类化石均采自关底组IV段下部(图3)。在近年的野外考察与化石发掘过程中,我们在关底组III段中也发现含鱼类大化石的数个层位(图3), 并采获一些化石材料,另在关底组I段下部之顶细砾岩中也发现少量鱼类微体化石,但限于时间和精力至今还没有开展相关的研究工作。曲靖周边地区关底组中潇湘脊椎动物群的发现与研究不仅为探索有颌类起源与早期分化提供了一个最重要的窗口,而且也为解决我国南方志留纪海相地层的划分与对比等地层学问题提供了可靠的古鱼类学证据(Zhao and Zhu, 2010, 2015)。
目前最新数据表明,关底组中的鱼化石主要赋存于关底组I, III及IV段之内,其产出的具体层位包括I段的中-下部、整个III段及IV段的下部(图3)。潇湘脊椎动物群的扬子鱼类组合主要产出于关底组III段及IV段的下部,而红庙鱼类组合则产出于妙高组和玉龙寺组的下部(图3)。
5 关底组的时代讨论
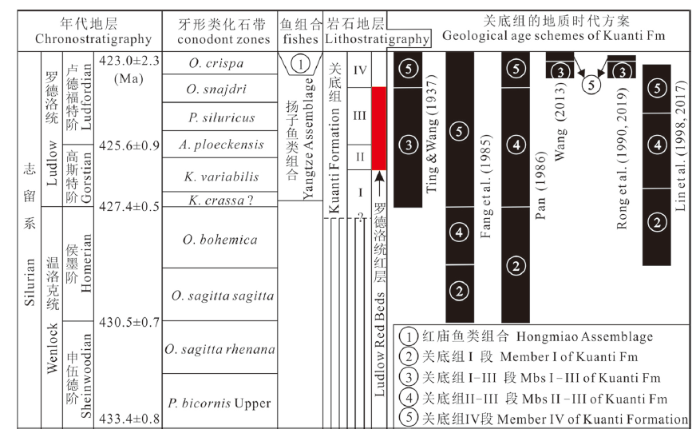
志留纪最常见的化石有笔石、腕足类和珊瑚等无脊椎动物,其中笔石是有效解决志留纪地层研究中一些重要地层学问题的第一主导化石门类,笔石带是志留系分统、分阶界线的主要依据。除笔石外,牙形类以及几丁虫(胞石)、疑源类、脊椎动物微体化石近年来也常被认为是在志留系生物地层的研究中十分重要的化石门类,其中牙形类更被认为是志留系第二主导化石门类(王成源,2005)。由于缺少深海相笔石化石,曲靖地区关底组的地质时代归属历来存在争议、没有定论。丁、王二氏(1937)最初将曲靖地区的关底组(仅相当本文关底组I-III段)划归上志留亚系(图4)。其后许多地层古生物****基于对无脊椎动物不同门类化石的研究,如Wang (1944, 1947), Sun and Wang (1946), 丁春鸣(1988)对珊瑚的研究,陈均远等(1981)对鹦鹉螺的研究,方润森等(1985)对介形虫的研究(图4), 将关底组划归温洛克统。潘江(1986)、Pan and Dineley (1988)通过对关底组I段(原称岳家山组)鱼化石的研究,认为关底组I段可与迴星哨组对比,应归入温洛克统,其上的关底组II-IV段(原称关底组+妙高组下段)均归入罗德洛统(图4)。戎嘉余等(1990, 2019)认为关底组(仅相当本文关底组I-III段)与迴星哨组不能对比,并将其划归罗德洛统上部(图4)。王尚启等(1992)根据对介形虫的研究也指出将关底组(仅相当本文关底组I-III段)归入温洛克统是不可靠的,并建议其时代为罗德洛世。何心一、陈建强(2000)依据珊瑚的研究成果认为,将岳家山段(相当于本文中的关底组I-II段)归入温洛克统、关底组上部即本文中的关底组III-IV段归入罗德洛统较为合理,但随后他们又指出,曲靖地区关底组的时代难以根据珊瑚群的时代确定为罗德洛世(何心一、陈建强,2004)。近40年来将关底组II-IV段归入罗德洛统的观点已陆续得到三叶虫、腕足类、胞石、牙形类等门类研究成果的支持。伍鸿基(1977)对三叶虫的详细研究,戎嘉余、杨学长(1978, 1981), 葛治洲等(1979)对腕足类的研究,王成源(1980, 1981, 1998, 2001, 2011, 2013)与Walliser and Wang (1989)对牙形类的研究,Geng et al. (1997)对胞石的研究,都支持将关底组(相当本文中关底组I-III段)归入罗德洛统(图4)。林宝玉等(2017)认为含牙形类Ozarkodina crispa的原岳家山组的上部应归入罗德洛统,而其下不含O. crispa的地层时代则应是温洛克世晚期(图4)。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4滇东关底组地质时代的代表性归属方案
Illustrations: ③+⑤=关底组+妙高组下部 Kuanti Formation (Fm) + the lower part of Miaokao Fm (Ting and Wang, 1937; 王成源,2013; 戎嘉余等,1990, 2019);②+④+⑤=岳家山组+关底组+妙高组下部 Yuejiashan Fm + Kuanti Fm + the lower part of Miaokao Fm (方润森等,1985; 潘江,1986; 林宝玉等,1998, 2017)
Fig. 4The representative schemes of geological age of the Kuanti Formation in East Yunnan
曲靖周边地区的志留系中含有大量的牙形类化石,特别是志留纪罗德洛世最晚期牙形类带化石Ozarkodina crispa在关底组、妙高组及玉龙寺组中的发现,为关底组地质时代的确定以及全球地层对比提供了重要的古生物学依据(王成源,1980, 1981, 1998, 2001, 2013; Walliser and Wang, 1989; 图4), 目前在没有笔石的关底组生物地层学研究中,牙形类无疑是确定关底组地质时代、提高国际对比精度的首选化石门类。然而,关于O. crispa的时代,金淳泰等(1992)曾有过质疑,推测该种可能时限较长、且在中国的出现可能比欧美早。另外目前国际上对O. crispa带的时代归属尤其是其上、下界的界定也不确定,至少有以下3种意见:1) 带化石O. crispa的消失是罗德洛世末的标志(Corradini and Corriga, 2012); 2) O. crispa可延至普里道利世(王成源, 2013; Corradini et al., 2015); 3) O. crispa在欧洲可下延到罗德洛世晚高斯特期(late Gorstian) (Viira and Aldridge, 1998)。从严格的意义上来讲,牙形类O. crispa带是志留系历程最短的一个带,在欧洲该带是介于O. snajdri带和O. remscheidensis eosteinhornensis带之间的一个罗德洛世最晚期的牙形类化石带,在波罗的海周边地区其所限地层厚仅数米至数十米(Viira and Aldridge, 1998), 而在曲靖周边地区含O. crispa化石的地层累加起来有1000多米厚,尽管已用曲靖地区在罗德洛世时快速沉降来予以解释(王成源,1980; Walliser and Wang, 1989), 但利用牙形类O. crispa的延限带来标定曲靖周边地区志留纪罗德洛世晚期的地层并不可取,这实际上与在欧洲牙形类O. crispa带通常与O. snajdri带和O. r. eosteinhornensis带存在互相重叠的事实也相矛盾(Viira and Aldridge, 1998)。曲靖地区的牙形类O. crispa非常丰富且类型多样,可划分出α, β, δ和γ 4个不同的形态型,该种的正型α类型延限时间最长,从关底组II段上部——王成源(2001)所提及的岳家山段上部[最低到丁春鸣(1988)所测剖面的12层,相当本文中所测剖面的15层]一直到玉龙寺组上部都有发现,而其他3种类型则主要见于关底组IV段中上部——Walliser and Wang (1989)所提及的妙高组下段,无疑关底组IV段中上部为其顶峰带。结合O. crispa在曲靖周边地区志留系中的这种分布特征,笔者认为在华南利用该种的顶峰带确定O. crispa带更为合理。基于此,曲靖地区含大量O. crispa的关底组IV段中上部无疑应归入罗德洛世最晚期O. crispa带;而含少量O. crispa的妙高组及玉龙寺组可归入普里道利统;另鉴于O. crispa在欧洲最低可下延到罗德洛世晚高斯特期(late Gorstian) (Viira and Aldridge, 1998), 本文认为关底组III-IV段下部的时代则可归入罗德洛世早-中卢德福特期(early to middle Ludfordian), 而含少量O. crispa的关底组II段上部及其下不含此牙形类化石的地层(包括关底组II段中下部及I段), 在没有发现确凿化石证据之前可暂归入罗德洛统高斯特阶较为稳妥,当然关底组I段的下限并不排除可下延至温洛克统的可能。上述方案也得到了近年来鱼类微体化石深入研究与分析的支持(图3)。
自国际地质对比计划(IGCP) 328项目实施以来,鱼类微体化石在志留、泥盆纪地层划分对比研究中所起到的重要作用开始被密切关注(Blieck and Turner, 2000; 戎嘉余、陈旭,2000)。 志留纪鱼类微体化石特别有助于解决缺少无脊椎动物标准化石的志留纪地层的精细划分与精确对比等重要的地层学问题(赵文金、朱敏,2014)。戎嘉余等(2019)指出,志留纪鱼类及其微体化石多产自近岸浅水海相沉积物中,以往常被忽视;近年来,志留纪鱼类的系统分类学、生物地层学研究越来越深入,精美的化石不断被发现,演化生物学和地层对比意义也逐渐显现。近十余年来,曲靖周边地区志留系中鱼类大化石及微体化石的研究为志留纪含鱼地层的时代确定与对比提供了可靠的证据。
根据曲靖周边地区志留系中鱼化石的类型及分布特征,Zhao and Zhu (2010)在潇湘脊椎动物群中识别出两个鱼类化石组合——分别赋存于关底组I段中部-IV段及妙高组-玉龙寺组底部之内的扬子组合与红庙组合(图3, 4), 其中后者主要依据硬骨鱼类及棘鱼类的鳞片材料所建立(赵文金、朱敏,2014), 产自妙高组下部的棘鱼类主要以棒棘鱼Gomphonchus, 背棘鱼Nostolepis和哈尼鱼Hanilepis三属为代表,硬骨鱼类则以纳西鱼Naxilepis和舌鳞鱼Ligulalepis为代表;而产自妙高组上部及玉龙寺组上易剥页岩之下地层中的棘鱼类主要以背棘鱼Nostolepis, 孔棘鱼Poracanthodes和哈尼鱼Hanilepis三属为代表,硬骨鱼类则以佧佤鱼Kawalepis为代表(图3)。潇湘脊椎动物群中具有时代意义的是红庙组合中的棘鱼类,在曲靖地区志留系中最丰富的Nostolepis是一类广泛分布于欧洲、北美、大洋洲及亚洲的栅棘鱼类,而Poracanthodes与Gomphonchus则是欧洲波罗的海周边地区常见的锉棘鱼类,它们的时限均主要限于罗德洛世-早泥盆世。近年来国内外同行对棘鱼类的研究表明,立陶宛东部地区罗德洛世-普里道利世的棘鱼类化石带中通常也见有在中国南方发现的三个棘鱼类属的分子,产出于玉龙寺组下部、上部的条纹背棘鱼Nostolepis striata更是普里道利世Monospina erecta化石带的代表分子(Valiukevi?ius, 2005)。据此,玉龙寺组可归入到普里道利统中上部,而其下红庙鱼类组合的主要赋存层位(包括妙高组及玉龙寺组底部)则可归入到普里道利统下部,这与关底组IV段中上部所含丰富的牙形类O. crispa所指示的地质时间并不矛盾。另在关底组III段上部除发现一些与IV段中盾皮鱼类相似的化石材料以外,也有硬骨鱼类纳西鱼Naxilepis鳞片的产出(王念忠、董致中,1989), 这表明关底组III段上部乃至整个III段归入罗德洛统上部卢德福特阶应无问题(图3, 4)。因此,依据鱼类化石资料,曲靖周边地区由扬子鱼类组合组成的潇湘脊椎动物群的主体产出层位——关底组III-IV段下部应归入罗德洛统卢德福特阶,而扬子鱼类组合在曲靖周边地区可下延至关底组I段下部之顶,相关的关底组I段地层及其上目前尚未见鱼类化石的关底组II段暂归入罗德洛统高斯特阶较为稳妥,关底组I段下部不含鱼类化石的地层则可暂时全部归入高斯特阶或者部分归入到温洛克统(图3, 4)。
综上所述,依据牙形类及鱼类微体化石,曲靖周边地区关底组的地质时代目前可以基本确定下来:其III-IV段的地层可归入到罗德洛统卢德福特阶,而I段中部-II段的地层则可归入到罗德洛统高斯特阶,I段下部地层目前还不能确定是全部归入到高斯特阶,还是部分可下延到温洛克统之中(图3, 4), 这有待深入的古生物学研究工作的开展。
6 结论
曲靖周边地区出露的关底组自创名以来备受关注,但其涵义及划分对比一直存有争议。笔者在实测一条较完整且连续的上铁路-东坡剖面的基础上,主要依据岩石学特征及丰富的鱼类化石资料,并结合牙形类、腕足类等无脊椎动物化石的研究进展,重新厘定了关底组的定义,划分出4个不同的次级岩石地层单元,综合分析讨论了关底组的地质时代,取得了对关底组如下的一些新认知:(1) 关底组是一套以细碎屑岩为主的滨浅海相红层沉积,在曲靖周边广泛出露于下寒武统龙王庙组或中寒武统双龙潭组之上、以浅灰色瘤状灰岩夹页岩为主的妙高组之下。
(2) 根据岩性变化特征将关底组划分为4段,从下往上依次为I段岳家山段、II段崇家湾段、III段彩莲段及IV段东坡段,曾从其下部划分出的岳家山组一名应予以废弃。
(3) 关底组的时限主要限于罗德洛世卢德福特期,其上限不会晚于牙形类Ozarkodina crispa带,其下限可能会下延到温洛克世,这有待于岳家山段下部不含化石层位当中化石的新发现。
致谢
张江永研究员与徐光辉研究员审阅文稿并提出了宝贵的修改意见,崔心东、李强、彭礼建参加了野外地质剖面的测制,在此一并致以谢意。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 8]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 4]
[本文引用: 3]
[本文引用: 6]
[本文引用: 4]
[本文引用: 3]
[本文引用: 1]
[本文引用: 3]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 6]
[本文引用: 1]
[本文引用: 4]
[本文引用: 5]
[本文引用: 4]
[本文引用: 3]
[本文引用: 3]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 5]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
DOIURLPMID [本文引用: 2]

The evolutionary history of osteichthyans (bony fishes plus tetrapods) extends back to the Ludlow epoch of the Silurian period. However, these Silurian forms have been documented exclusively by fragmentary fossils. Here we report the discovery of an exceptionally preserved primitive fish from the Ludlow of Yunnan, China, that represents the oldest near-complete gnathostome (jawed vertebrate). The postcranial skeleton of this fish includes a primitive pectoral girdle and median fin spine as in non-osteichthyan gnathostomes, but a derived macromeric squamation as in crown osteichthyans, and substantiates the unexpected mix of postcranial features in basal sarcopterygians, previously restored from the disarticulated remains of Psarolepis. As the oldest articulated sarcopterygian, the new taxon offers insights into the origin and early divergence of osteichthyans, and indicates that the minimum date for the actinopterygian-sarcopterygian split was no later than 419 million years ago.
[本文引用: 2]
DOIURLPMID [本文引用: 2]
The gnathostome (jawed vertebrate) crown group comprises two extant clades with contrasting character complements. Notably, Chondrichthyes (cartilaginous fish) lack the large dermal bones that characterize Osteichthyes (bony fish and tetrapods). The polarities of these differences, and the morphology of the last common ancestor of crown gnathostomes, are the subject of continuing debate. Here we describe a three-dimensionally preserved 419-million-year-old placoderm fish from the Silurian of China that represents the first stem gnathostome with dermal marginal jaw bones (premaxilla, maxilla and dentary), features previously restricted to Osteichthyes. A phylogenetic analysis places the new form near the top of the gnathostome stem group but does not fully resolve its relationships to other placoderms. The analysis also assigns all acanthodians to the chondrichthyan stem group. These results suggest that the last common ancestor of Chondrichthyes and Osteichthyes had a macromeric dermal skeleton, and provide a new framework for studying crown gnathostome divergence.
DOIURLPMID [本文引用: 2]
The discovery of Entelognathus revealed the presence of maxilla, premaxilla, and dentary, supposedly diagnostic osteichthyan bones, in a Silurian placoderm. However, the relationship between these marginal jaw bones and the gnathal plates of conventional placoderms, thought to represent the inner dental arcade, remains uncertain. Here we report a second Silurian maxillate placoderm, which bridges the gnathal and maxillate conditions. We propose that the maxilla, premaxilla, and dentary are homologous to the gnathal plates of placoderms and that all belong to the same dental arcade. The gnathal-maxillate transformation occurred concurrently in upper and lower jaws, predating the addition of infradentary bones to the lower jaw.
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}