 ,1,2, 张晓晓1,2,3,4, 李春晓1,2,3
,1,2, 张晓晓1,2,3,4, 李春晓1,2,3Reappraisal of Serridentinus gobiensis Osborn & Granger and Miomastodon tongxinensis Chen: the validity of Miomastodon
WANG Shi-Qi,1,2, ZHANG Xiao-Xiao1,2,3,4, LI Chun-Xiao1,2,3收稿日期:2019-09-26网络出版日期:2020-04-20
| 基金资助: |
Received:2019-09-26Online:2020-04-20
作者简介 About authors
wangshiqi@ivpp.ac.cn

摘要
中国北方中中新世的乳齿象类戈壁锯齿象(Serridentinus gobiensis
关键词:
Abstract
The elephantimorph proboscideans, Serridentinus gobiensis
Keywords:
PDF (3911KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王世骐, 张晓晓, 李春晓. 戈壁锯齿象(Serridentinus gobiensis Osborn & Granger, 1932)和同心中新乳齿象(Miomastodon tongxinensis Chen, 1978)再研究:关于粗壮型轭齿象(Zygolophodon)的讨论. 古脊椎动物学报[J], 2020, 58(2): 134-158 DOI:10.19615/j.cnki.1000-3118.200310
WANG Shi-Qi, ZHANG Xiao-Xiao, LI Chun-Xiao.
Serridentinus Osborn, 1923, was established based on Mastodon productus Cope, 1875, and several North American Mastodon species. Later, Osborn and his colleagues expanded the extent of Serridentinus, and published several Eurasian species, e.g., S. gobiensis Osborn & Granger, 1932, from Tunggur, northern China. The holotype is a right hemimandible carrying the moderately to deeply worn m2 and the erupting m3, with complete mandibular symphysis and lower tusks. Miomastodon Osborn, 1922, was established mainly based on Mastodon merriami Osborn, 1921, which he regarded as the first “true Mastodon” in North America. He also broadened the name Miomastodon to Eurasian species, e.g., Mio. depereti Osborn, 1936. Chen (1978) reported another Eurasian species, Mio. tongxinensis from Tongxin, northern China, which is represented by a pair of deeply worn lower and upper third molars.
Many debates occurred over the validity of Serridentinus gobiensis and Miomastodon tongxinensis subsequently. Tobien (1972) did not agree with the validity of the genus Serridentinus, and synonymized most of its species with Gomphotherium. However, he considered that the molar of S. gobiensis displays the zygodont pattern (Tobien, 1972). Tassy (1985) thought that S. gobiensis belongs to the robust type of Zygolophodon. Later, Tobien et al. (1988) attributed all the Chinese samples from Middle or early Late Miocene with more or less zygodont features into the single species, Z. gobiensis, including Mio. tongxinensis. Tobien (1996) further considered that Z. gobiensis belongs to the Z. turicensis group. However, Z. gobiensis sensu Tobien et al. (1988) bears much variation and spans quite a long geological period. Therefore, further study of Z. gobiensis sensu Tobien et al. (1988) should be carried out, and the key problem is the taxonomy of S. gobiensis.
1 Material and methods
1.1 Institutional abbreviations
AMNH and FAMNH, American Museum of Natural History, New York, USA; HMV, Hezheng Paleozoological Museum, Hezheng, China; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; MNHN, Muséum National d'Histoire Naturelle, Paris, France; NHMW, Naturhistorisches Museum Wien, Vienna, Austria; NMB, Naturhistorisches Museum Basel, Basel, Switzerland; SNSB-BSPG, Staatliche Naturwissenschaftliche Sammlungen Bayerns - Bayerische Staatssammlung für Pal?ontologie und Geologie, Munich, Germany; THP, Tianjin Natural History Museum, Tianjin, China.1.2 Materials and terminology
The material described in the present work is housed in AMNH and IVPP. The comparative material is housed in AMNH, IVPP, HMV, MNHN, NHMW, NMB, SNSB-BSPG and THP. Others were illustrated in previous publications (Cope, 1873; Lortet and Chantre, 1878; Schlesinger, 1917; Osborn, 1921, 1922,1929,1936; Borissiak, 1936; Lehmann, 1950; Tassy, 1977, 2013, 2014; Madden and Storer, 1985; Chen, 1988; G?hlich, 2010; Lofgren and Anand, 2011; Wang et al., 2017).The terminology of the occlusal structure of gomphotheriid molars and mandible follows Tassy (2013, 2014; Fig. 1A) with several modifications. Specifically, we used the terms pretrite central conule and pretrite crescentoid for different crown elements, which Osborn (1936) had already dealt with (he used conule or serration for pretrite central conule and spur or crest for crescentoid, see Osborn, 1936:393). A pretrite crescentoid (green color) is a thick or thin enamel projection that originates from the mesial or distal side of the pretrite main cusp(id) and runs to the base of the loph(id)s (Fig. 1D, green color). The proximal end of a crescentoid merges closely with the attached main cusp(id) without a sulcus between them. Whereas, a pretrite central conule (blue color) is a round, bulbous cone (Fig. 1E, blue color), or subdivided enamel cones that are either arranged in a line or serrated (Fig. 1A, posterior pretrite central conule 2). It should be noted that the boundary between crescentoids and central conule might be vague after moderate or deep wear. Pretrite central conules rise directedly from the base of the interloph(id) or the anterior cingulum(id), with a sulcus that clearly separates the adjacent loph(id). Pretrite crescentoids are lacking in the most typical bunodonts (i.e., Gomphotherium angustidens) and pretrite central conule are absent in typical zygodonts (i.e., Mammut borsoni). However, in some cases, the two elements coexist (Fig. 1A). A short and thick pretrite crescentoid derives from the peak of the main cusp(id), it is followed by a single or subdivided pretrite central conule(s) that is separated from the pretrite crescentoid by a sulcus, e.g., in Gomphotherium productum.
Fig. 1
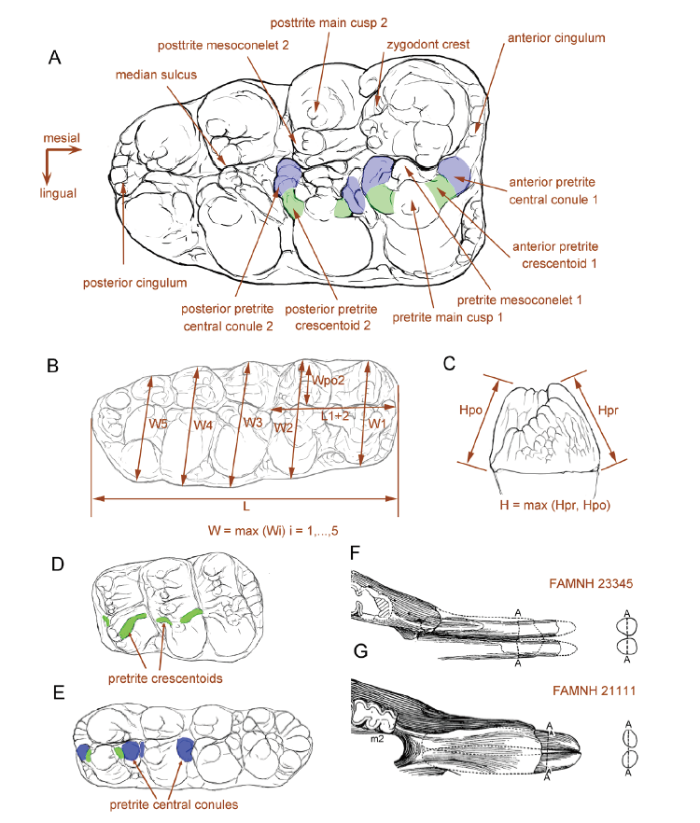 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Terminology and measurements of mastodont molars
A. right M3 of Sinomastodon praeintermedius, denoting the terminology of tooth crown; B. molar crown measurements; C. molar height measurements; D, E. the development of pretrite accessory elements in a typical zygodont molar, Zygolophodon turicensis, only pretrite crescentoids present (D), and in a typical bunodont molar, Gomphotherium connexum, only pretrite central conule present in the lophid 2 (E);F, G. the cross-sections of lower tusks, showing the “erected oval cross-sectioned mandibular tusk” in Miomastodon merriami, in which Ddv > Dml (F), and the “laid oval cross-sectioned” in Gomphotherium productum, in which Ddv < Dml (G). Note that the discrimination of the pretrite accessory elements in the panels A, D, E: green color, pretrite crescentoids; blue color, pretrite central conules. Panels F and G were modified from Frick (1933). Abbreviations: L. length; L1+2. length of the first two loph(id)s; H. height;Hpo. height of the posttrite side; Hpr. height of the pretrite side; W. width;W1, 2, …, 5. width of the 1st, 2nd, …, 5th loph(id); Wpo2. width of the second posttrite half loph(id)
1.3 Measurements, specimen illustrations, and data analyses
Mandibular and cheek teeth measurements follow Tassy (2013,2014; Fig. 1B, C). Measurements were preferentially obtained using callipers (in mm). If not available, photos and 3D digital models were also used, and the data were obtained using the public software Image J (V 1.48) (Schneider et al., 2012).Illustrations of mandibles and molars in the present article were based on photos taken by a Nikon D7 100 camera equipped with an AF-S NIKKOR 18-300 mm optical zoom lenses, or snapshots of 3D digital models generated by a handheld Artec Spider 3-dimensional scanner.
Zygodont taxa are characterized by the remarkable widening of their cheek teeth, for more efficient bolus compression (K?nigswald, 2016), particularly by the widening of the posttrite half loph(id)s. Thus, in our cheek teeth measurement protocol, in addition to the general measurements as described in Tassy (2014), we measured the posttrite width of the second loph(id), from the median sulcus to the summit of the posttrite main cusp(id) (Wpo2; Fig. 1B). For comparison of biometric data between taxa with different size, the raw data should be normalized. For the posttrite width of the second loph(id) (Wpo2), we used the width of the second loph(id) (W2) for normalization, i.e. Wpo2/W2 (Fig. 1B). For maximal width (W), we measured the length of the first two loph(id)s (L1+2) rather than the whole length for normalization, i.e. W/L1+2. This treatment is advantageous for comparing M1-M3 or m1-m3 in the same plot. In the present paper, we only compared the lower cheek teeth because the upper cheek teeth are very rare in our concerning material. We figured both width vs. length of m3 and Wpo2/W2 vs. W/L1+2.
For the mandibular tusk measurements, we measured the dorsoventral (Ddv) and mediolateral diameters (Dml), and figured Dml vs. Ddv for the mandibular tusk comparison. For comparing the mandibular symphysis, we measured the length of the symphysis (Ls), maximal width of the symphysis (Ws), and length of the cheek tooth row (Lt) (after Tassy, 2013, measurements 2, 11, and 24, respectively). We figured Ls/Lt vs. Ws/Lt, which means the relative length and relative width of mandibular symphysis, respectively.
2 Systematic paleontology
Order Proboscidea Illiger, 1811Family Mammutidae Hay, 1922
Genus MiomastodonOsborn, 1922
Type speciesMiomastodon merriami ( Osborn, 1921).
Referred speciesMiomastodon tongxinensis Chen, 1978, Miomastodon gobiensis (Osborn & Granger, 1932), Miomastodon metachinjiensis (Osborn, 1929).
Diagnosis Longirostrine trilophodont elephantimorph with narrow and moderately elongated symphysis; mandibular tusks straight, parallel-protruding, close to each other, and with an “erected oval cross-section” (Ddv>Dml); posttrite half lophids moderately subdivided, mesiodistally compressed, and lacking posttrite central conules; pretrite crescentoids thicker than those of Zygolophodon turicensis, especially for lower molars; pretrite central conules usually present on the first and second interlophids of lower molars.
Differs from the other members of the Mammutidae (Losodokodon, Eozygodon, Zygolophodon, and Mammut, Sinomammut) in the relatively bunodont characters of the cheek teeth, in the slightly chevroned and alternatively positioned half lophid of m3; also differs from Mammut in the relatively longer symphysis (Ls/Lt>1, see Fig. 5D and Table S3) and mandibular tusks. Differs from Gomphotherium subtapiroideum and G. tassyi in weaker mandibular symphysis (Ws/Lt<0.4, see Fig. 5D and Table S3); in mandibular tusks with oval rather than pyriform cross-section. Differs from the other longirostrine trilophodont gomphotheriid taxa in the more zygodont characters of the cheek teeth.
Fig. 5
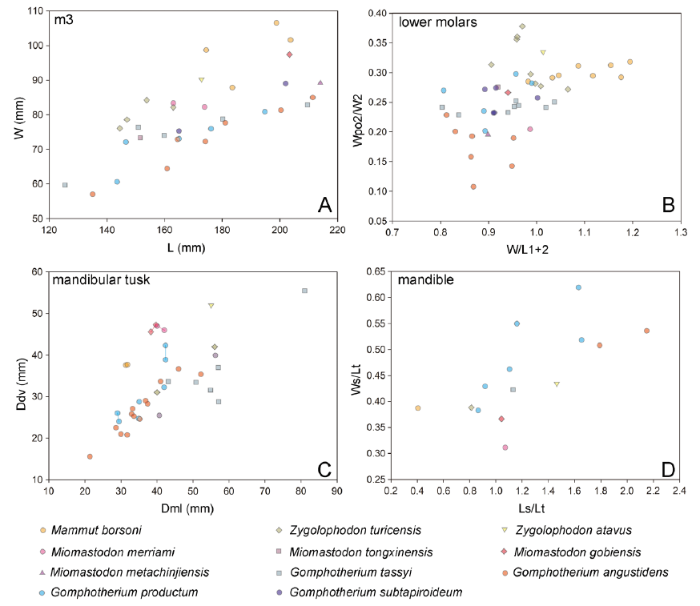 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Scatter plots of bunodont and zygodont cheek teeth and mandibular tusks of the relevant taxa
A. m3 length vs. width (W/L); B. normalized maximal width (W/L1+2) vs. normalized posttrite width of lophid 2 (Wpo2/W2) for lower molars; C. cross-sectional mediolateral diameter (Dml) vs. dorsoventral (Ddv) diameter of mandibular tusk; D. mandibular measurements, the relative mandibular length (Ls/Lt) vs. the relative mandibular width (Ws/Lt). The raw data see Tables S1-S3
Miomastodon gobiensis (Osborn & Granger, 1932)
(Figs. 2A-D, 4A-C; Tables 1, 2)
Serridentinus gobiensis Osborn and Granger, 1932
Zygolophodon gobiensis (Osborn & Granger, 1932) Tobien, 1972, p. 176
Zygolophodon gobiensis (Osborn & Granger, 1932) Tassy, 1985, p. 511
partim Zygolophodon gobiensis (Osborn & Granger, 1932) Tobien et al., 1988, p. 146-156, figs. 39, 41, 43, 44
non Zygolophodon nemonguensis Chow and Chang, 1961
non Zygolophodon gromovae Dubrovo, 1970
non Zygolophodon jiningensis Chow and Chang, 1974
non Zygolophodon chinjiensis (Osborn, 1929) Chow et al., 1978
Type specimen AMNH 26461, a right hemimandible carrying mandibular symphysis with a pair of lower tusks. The tooth row consists of the vacancy of m1, moderately worn m2, and erupting m3.
Type locality and horizon 6.4 km (4 miles) northwest to the Wolf Camp (Zhunwuguer locality), Tunggur Formation, Tamuqin Fauna, ~MN7/8, see below (Osborn and Granger, 1932; Wang et al., 2003; Qiu et al., 2013).
Occurrence Tunggur region, Nei Mongol, China, late Middle Miocene.
Referred material AMNH 26476, a right m3 with partial mandibular fragments (see Tobien et al., 1988:fig. 41).
Differential diagnoses Miomastodon with relatively large size (~120% m3 length of Mio. merriami) and relatively longer symphysis. Differs from Mio. merriami in the longer mandibular symphysis and in possessing the pretrite central conules in the first and second interlophids of the lower molars; differs from Mio. tongxinensis in the larger dimensions and in the narrower m3 with a strong fourth lophid; differs from Mio. metachinjiensis in the relatively stronger pretrite central conules, in the greater subdivision of the posttrite half lophids, and in the smaller m3 dimensions.
Remarks The type hemimandible of Miomastodon gobiensisOsborn & Granger, 1932 was discovered by Andrews, the head of the Central Asiatic Expedition 1930, from an isolated hill north of the Wolf Camp (Osborn and Granger, 1932), Middle Miocene Tunggur Formation. In recent publications, this locality was recognized as “Zhunwuguer” (Wang et al., 2003). Based on Spock (1929), Mio. gobiensis was recovered from the upmost cross-bedded sandstones which overlies a thick layer of limestones. It possibly coexisted with a very derived Platybelodon (more derived than the type species of Pl. grangeri), and the layer also produces the Tamuqing small mammal assemblage, which is the youngest small mammal assemblage in the Tunggur Formation, with the approximate age of 11.8 Ma (Wang et al., 2003; Qiu et al., 2013).
Descriptions (see Osborn and Granger, 1932:11; Tobien et al., 1988:147, 149) In AMNH 26461 (Fig. 2), the left corpus and ramus are broken while the other parts are complete. In dorsal view (Fig. 2A) the ramus is thin and plate-like. The condyloid process of the mandible is oval and its long axis is transversely oriented. The shape of mandibular corpus looks like a triangle of which the acute angle is anteriorly oriented. The m1 alveolus was healing, leaving a relatively large vacancy. The mid-axes of the m2 and m3 are not in line. The symphysis is moderately elongated. It is relatively narrow and the distal part is only slightly widened. The symphyseal trough is deep and the two interalveolar crests border each side of the symphyseal trough. The posterior end of the symphysis is almost in line with the anterior end of the m1 alveolus. The distal end of the symphysis displays an obtuse angle with an anteriorly oriented apex. In lateral view (Fig. 2B), the ramus is high, and the dorsally protruded condyloid process is much higher than the hook-like coronoid process. These two processes are linked by a smoothly curved mandibular notch. The masseter fossa is trapezoidal. The angular process is rather blunt, but slightly ventrally bulging. The anterior end of the mandibular corpus is slightly thicker than its posterior end. The symphysis stretches slightly ventrally, and the ventral border of the mandible is nearly straight. The anterior mental foramen is large, and is distant from the tooth row, but the posterior mental foramina are absent. In rostral view (Fig. 2C), the symphyseal trough is smooth and deep. It dips down rostrally and shows two medially oblique edges at the distal end. The anterior mental foramen is relatively rounded and faces anteriorly. The ascending ramus is high and thin, and the large mandibular condyle is slightly medially oblique.
Fig. 2
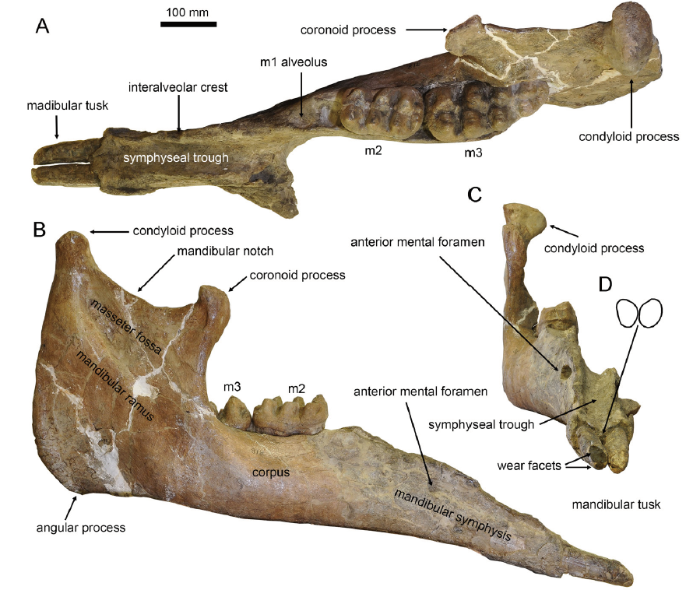 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Type mandible of Miomastodon gobiensis, AMNH 26461, from the Zhunwuguer locality, Tunggur region, Nei Mongol, in dorsal (A), lateral (B), and rostral (C) views, and the contour of the cross-section of mandibular tusks, in rostral view (D)
The mandibular tusks (Fig. 2A-D) are simply rod-like. They are straight, closely apposed, with a relatively short exposed length (~130 mm, precise measurements see Table 1). The cross-section is oval and its maximal axis is slightly dorsolateral-ventromedial oriented. The dorsoventral diameter is larger than the mediolateral diameter, which we call the “erected oval cross-section” (the ratio of Ddv/Dml > 1, see Table S2, Fig. 2D). The main wear facet (Fig. 2C) is also oval shaped. It obliquely cuts the rostrodorsal end of the tusk. Another small facet (Fig. 2C) is just beneath the main facet. It is perpendicular to the longitudinal axis of the tooth.
Table 1
Table 1Mandibular and tusk measurements of Miomastodon gobiensis* (mm)
| maximum length | 1190 | |
|---|---|---|
| symphyseal length | 338.41 | |
| alveolar distance (from the most salient point of the retromolar trigonum to the symphyseal border of the corpus) | 424.1 | |
| ventral length measured from the gonion (angulus mandibular) to the tip of the symphysis | 965 | |
| width of corpus measured at the root of the ramus | 148.52 | |
| width of corpus measured at the anterioralveolus (or the grinding tooth if the alveolus is entirely resorbed) | 62.01 | |
| anterior symphyseal width | 113.75 | |
| maximum symphyseal width in the anterior part of the symphysis | 118.09 | |
| minimum symphyseal width in the posterior part of the symphysis | 69.88×2 | |
| maximum width of rostral trough in the anterior part of the symphysis | 80.17 | |
| minimum width of rostral trough in the posterior part of the symphysis | 82.76 | |
| rostral height measured at the symphyseal border (measurement taken perpendicular to the ventral border of the symphyseal rostrum) | 146.84 | |
| rostral height measured at the tip of rostrum (measurement as above) | 70.13 | |
| maximum mandibular height measured at the condyle perpendicular to the ventral border of the corpus | 467.97 | |
| maximum depth of ramus | 317.14 | |
| depth between gonion and coronoid processes | 364.98 | |
| height between gonion and condyle | 329.08 | |
| mid-alveolar length measured on the buccal side between the anterior alveolus (or grinding tooth if the alveolus is resorbed) and the root of the ramus | 321.73 | |
| exposed length of mandibular tusk | left | 134.25 |
| right | 131.5 | |
| width of mandibular tusk at the alveolu | left | 38.26 |
| right | 39.74 | |
| height of mandibular tusk at the alveolus | left | 45.60 |
| right | 47.32 | |
| length of wear facet of mandibular tusk | left | 35.33 |
| right | 48.31 |
新窗口打开|下载CSV
The m2 is moderately worn (Fig. 3A-C). In occlusal view, the width of lophids increases from anterior to posterior. The first pretrite half lophid is trifoliate with a small mesoconelet and nearly symmetrical anterior and posterior crescentoids. The crescentoids are thick, and a small and bulbous posterior central conule is present. The first posttrite half lophid is slightly mesiodistally compressed and the sulcus between the main cusp and the mesoconelets is clear. The first interlophid is relatively mesiodistally wide and is blocked by the pretrite central conules. A large conule is present at the entoflexid. The second lophid is similar to the first one. The anterior and posterior crescentoids are smaller than those of the first lophid. The second posterior pretrite central conule is also present. The posttrite half lophid displays a tendency for subdivision and mesiodistal compression, and the separation of the main cuspid and the mesoconelet can be observed. The second interlophid is also mesiodistally wide. The third pretrite half lophid possesses a small anterior pretrite crescentoid and a small anterior pretrite central conule, but the posterior pretrite crescentoid is unclear. The posttrite half lophid is more strongly subdivided than the anterior two, showing a series of serrated conelets, and the separation of the main cuspid and the mesoconelet cannot be observed. Cingulids are present on the anterior and posterior margins of the tooth and two large conelets arise on the center of the posterior cingulid. In buccal view (Fig. 3B), the lophids are “?-shaped”. The first interlophid is “U-shaped” and the second interlophid is “V-shaped”. The two interlophids are stuffed up by the central conules and crescentoids due to deep wear. In lingual view (Fig. 3C), the lophids are “Λ-shaped” and the interlophids are “V-shaped”.
Fig. 3
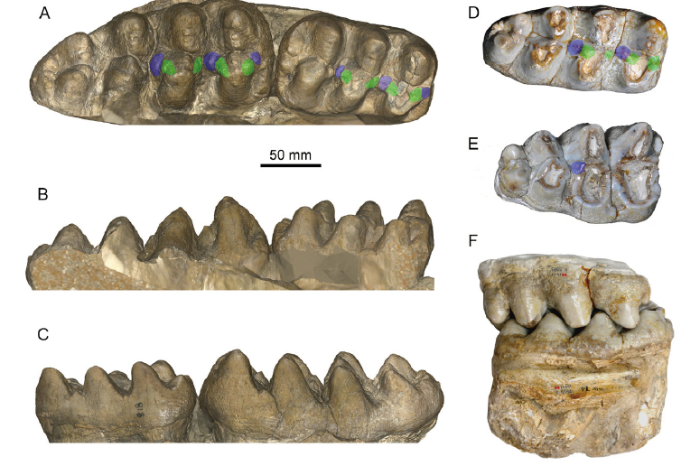 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Cheek teeth of Miomastodon gobiensis and Mio. tongxinensis
A-C. Mio. gobiensis, AMNH 26461, right m2 and m3 of the type specimen, from the Zhunwuguer locality, Tunggur region, Nei Mongol, China, the ramus has been removed in the 3D digital model, in occlusal (A), buccal (B), and lingual (C) views; D-F. Mio. tongxinensis, from Tongxin region, Ningxia, China: IVPP V 5584, the type right m3, in occlusal view (D), V 5585, the referred right M3, in occlusal view (E), and the occlusion of V 5584 and V 5585, in buccal view (F), indicating that they belong to the same individual+Note: green color, pretrite crescentoids; blue color, pretrite central conules
The m3 (Fig. 3A-C) is erupting from the first three lophids. However, the covering boney plate has been removed and the entire tooth is exposed. In occlusal view (Fig. 3A), this tooth has four lophids plus a strong posterior cingulid and is long leaf-shaped. Lophids 2 and 3 display a slight tendency for alternative position of half lophids and the posterior three lophids are slightly chevroned. The first pretrite half lophids has weak but subdivided mesoconelets. The pretrite anterior and posterior crescentoids are strong, and the anterior and posterior pretrite central conule rises from the anterior cingulid and the first interlophid, respectively. The first posttrite half lophid has a strong main cuspid and a subdivided mesoconelet. The anterior zygodont crest is present and the posterior one is absent. The second pretrite half lophid also has a small and subdivided mesoconelet. The anterior and posterior pretrite crescentoids are weak, the anterior pretrite central conule is almost absent, and the posterior one is relatively strong. The posttrite half lophid is composed of a line of small conelets, and the main cuspid and the mesoconelet are indistinguishable. The third pretrite half lophid possesses a main cuspid that is equivalent to the mesoconelet, the anterior pretrite crescentoid is very weak, and the other accessory structures are absent. The third posttrite half lophid is composed of four equal-sized conelets, and the main cuspid and the mesoconelets are indistinguishable. The third interlophid is mesiodistally open. The fourth lophid is almost the same as the third one, but much smaller. Cingulids are present on the anterior and posterior ends of the teeth. In buccal view (Fig. 3B), the lophids are “?-shaped”. The first interlophid is “U-shaped” and the second and third interlophids are “V-shaped”. The anterior two interlophids are half stuffed up by the central conules and crescentoids (central conules reach the half height of the interlophids). In lingual view (Fig. 3C), like the m2, the lophids are “Λ-shaped” and the interlophids are “V-shape”.
Miomastodon tongxinensis Chen, 1978
(Fig. 3D-F; Table 2)
Table 2
Table 2Cheek tooth measurements of Miomastodon* (mm)
| Miomastodon gobiensis | Miomastodon tongxinensis | |||||
|---|---|---|---|---|---|---|
| specimen | AMNH 26461 | AMNH 24676 | IVPP V 5584 | IVPP V 5585 | ||
| locus | right m2 | right m3 | right m3 | right m3 | right M3 | |
| L | 136.8 | 203.31 | 158.3 | 152.0 | 137.5 | |
| W1 | 75.19 | 91.29 | 74.65 | 69.0 | 81 | |
| W2 | 78.2 | ca.98 | 77.61 | 73.4 | 82 | |
| W3 | 87.52 | 75.51 | 67.6 | 66.5 | ||
| W4 | 63.34 | 51.5 | 44 | |||
| Hpo | 51.55+ | 68 | 42.89+ | 48.5+ | 48.5+ | |
新窗口打开|下载CSV
partim Zygolophodon gobiensis (Osborn & Granger, 1932) Tobien et al., 1988, p. 146-156, figs. 39, 41, 43, 44
Type specimen IVPP V 5584, a deeply worn right m3 (Chen, 1978:pl. II, fig. 2).
Type locality and horizon Zhangenbao Formation in the Tongxin region, Ningxia. Late Early or early Middle Miocene.
Referred specimen IVPP V 5585, a deeply worn right M3 (Chen, 1978:pl. II, fig. 1), probably belonging to the same individual as the type m3
Differential diagnosesMiomastodon with small dimensions (~85% m3 length of Mio. merriami and ~75% of Mio. gobiensis and Mio. metachinjiensis, precise measurements see Table 2). Differs from the other Miomastodon species in the smaller size, mostly because of the less developed fourth loph(id); also differs from Mio. merriami in the presence of the first and second posterior pretrite central conules of the lower molars.
Remarks Chen (1978) reported Miomastodon tongxinensis, represented by a pair of lower and upper third molars that are deeply worn. The size is fairly small, therefore might represent a senile female individual. The material was collected from Gujiazhuangzi, a local village of Tongxin region, where the Early to Middle Miocene Zhangenbao Formation is well exposed and produces abundant fossil mammals. The Zhangenbao Formation consists of five fluviolacustrine sedimental circles, and the first and second circles represent the late Early Miocene and the third to fifth represent the early Middle Miocene. The precise locality of Mio. tongxinensis is unclear, and there are two possibilities: one is from the second circle at Miaoerling, which is characterized by the ferrygineous sandstones attached on the specimens; and another one is from the fourth circle at Shataigou, the nearest fossil locality to the Gujiazhuangzi village.
Descriptions (see Chen, 1978:103, 104; Tobien et al., 1988:152, 153) IVPP V 5584 is a deeply worn right m3 (Fig. 3D). In occlusal view, the tooth is long leaf-shaped and is relatively narrow. The second lophid is slightly wider than the other lophids. The lophids 2 and 3 display a slight tendency for alternative position of half lophids and the posterior three lophids are slightly chevroned. The first pretrite half lophid is trifoliate. The enamel walls of the posterior pretrite crescentoid and central conule are connected to each other, only showing a distal inflated enamel loop. The posttrite half lophid shows mesiodistally compression and subdivision. The main cuspid is strongly oblique. Fairly weak anterior and posterior zygodont crests are present. The first interlophid is mesiodistally wide. The second pretrite half lophid is similar to the first one. The posterior pretrite central conule is strong, linked to the pretrite lophid by the thick posterior pretrite crescentoid. The second posttrite half lophid is also mesiodistally compressed, but it is more deeply worn than the first half lophid. The second interlophid is also mesiodistally wide. The anterior and posterior crescentoids of the third pretrite half lophid are weak, and only a small posterior pretrite central conule is present. The posttrite half lophid shows distinct separation of the main cuspid and the mesoconelet. The fourth pretrite half lophid has a strong mesoconelet that is anteriorly positioned and the posttrite half lophid is small. Cingulids are present on the anterior and posterior margins of the tooth.
IVPP V 5585 is a deeply worn right M3 (Fig. 3E) that very possibly belongs to the same individual as V 5584, as they are perfectly matched (Fig. 3F). In occlusal view, it is wide with a triangular shape. The posterior two lophs are chevroned. The first two pretrite half lophs are so deeply worn showing two oval or subcircular enamel rings. However, the posterior pretrite central conule of the second loph is prominent. The first two posttrite half lophs are mesiodistally compressed, and form two transversely elongated enamel rings. The anterior two interlophs are mesiodistally narrow, relative to those of the m3. On the third loph, the pretrite half loph lacks a posterior crescentoid and a central conule, and the posttrite half loph shows a clear groove between the main cusp and the mesoconelet. The fourth loph is very small, but the pretrite half loph possesses an anterior central conule and a mesoconelets. The posterior cingulum is almost absent.
3 Results and discussion
3.1 Molar comparisons
3.1.1 Comparison to Zygolophodon turicensisTobien et al. (1988) allocated most of the specimens from the middle to early Late Miocene of China showing less or more zygodont characters into one species, Zygolophodon gobiensis, including Serridentinus gobiensis Osborn & Granger, 1932, and Miomastodon tongxinensis Chen, 1978. Tobien et al. (1988:155) stated that the type mandible of Z. gobiensis (i.e., S. gobiensisOsborn & Granger, 1932) belongs to the robust or primitive morph-type. Other specimens, e.g., Z. gromovae Dubrovo, 1970, Z. (Turicius) nemonguensis Chow & Chang, 1961, and Z. jiningensis Chow & Chang, 1974, were attributed to the gracile or advanced morph-type of Z. gobiensis (Tobien et al., 1988). The two morph-types were also recognized in the European sample of Z. turicensis (Tobien, 1975; Tassy, 1985). The gracile type of Zygolophodon is indisputable, as the type specimen of Z. turicensis is absolutely the gracile type (see below). However, the robust type of Zygolophodon was always confused with various bunodont taxa, and thus gave rise to much controversy. Tobien et al. (1988) interpreted the two morph-types as a result of functional partition. However, the morphological distinction between the two types is clearly shown on the unworn specimens, and functional partition in a single species is untenable.
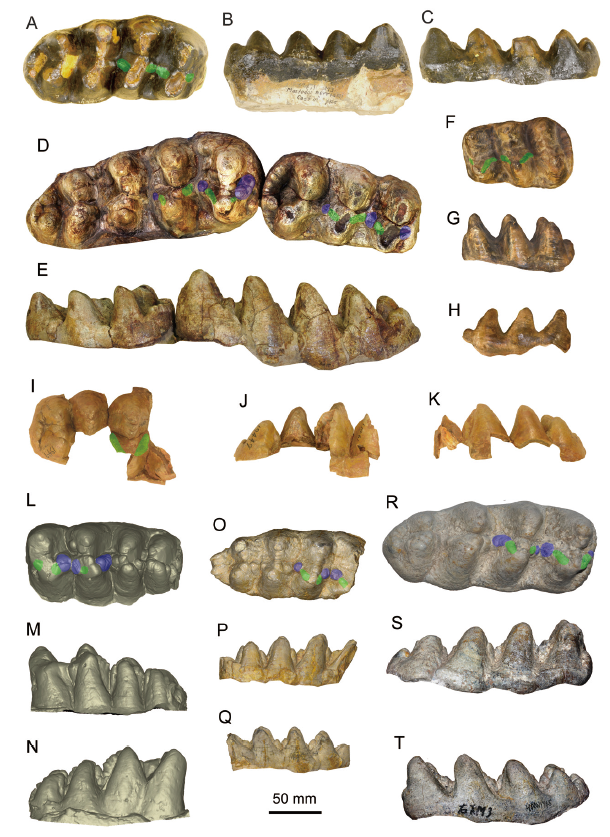
The type specimen of Zygolophodon turicensis is a left m2 from Elgg, Switzerland, Middle Miocene (Fig. 4F-H). It shows a high grade of zygodonty. In occlusal view (Fig. 4F), the posttrite half lophids are highly compressed; and the conelets are strongly subdivided, arranging in line as a sharp edge. The anterior and posterior crescentoids are sharp and slender, and no anterior and posterior central conules rise from the interlophids. In buccal and lingual views (Fig. 4G, H), the lophids are Λ-shaped preserving a sharp peak. In buccal view (Fig. 4G), the interlophids exhibit high and deep V-shaped notches. These features are typical in the gracile type of Z. turicensis.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Molars of the comparative species of the relevant taxa
A-C. Miomastodon merriami, AMNH 14471 (type cast), the right m3, from Thousand Creek Formation, Nevada, USA, in occlusal (A), buccal (B), and lingual (C) views; D, E. Mio. metachinjiensis, AMNH 19414 (type specimen), the right m2 and m3, from Chinji Bungalow, Pakistan, in occlusal (D) and lingual (E) views; F-H. Zygolophodon turicensis, NMB OSM1288 (type cast), the left m2, from Elgg, Kanton Zürich, Switzerland, in occlusal (F), buccal (G), and lingual (H) views; I-K. Z. chinjiensis, AMNH 19447 (type specimen), the right M3, from Chinji Bungalow, Pakistan, in occlusal (I), lingual (J), and buccal (K) views; L-N. Gomphotherium productum, AMNH 14383 (type cast), the left m3, from Santa Fé, New Mexico, USA, in occlusal (L), buccal (M), and lingual (N) views; O-Q. G. subtapiroideum, SNSB-BSPG 1959 II 11381, the right m3, from Sandelzhausen, Germany, in occlusal (O), buccal (P), and lingual (Q) views; R-T. G. tassyi, HMV 1945, the right m3, from Linxia Basin, China, in occlusal (R), buccal (S), and lingual (T) views+Note: green color, pretrite crescentoids; blue color, pretrite central conules
The molars of the type specimen of Serridentinus gobiensis, as the other robust type of Zygolophodon, display more bunodont morphology than Z. turicensis. In occlusal view (Fig. 3A), the posttrite half lophids are moderately subdivided, the separation of the posttrite main cuspid and the corresponding mesoconelet is unclear, and these arranged posttrite conelets are relatively blunt rather than sharp. The pretrite mesoconelets are more bulbous than those of the typical Z. turicensis. The anterior and posterior crescentoids are thicker than those of the typical Z. turicensis. More importantly, in the lophids 1 and 2, a round and bulbous posterior pretrite central conule is present at the distal end of the corresponding posterior pretrite crescentoid. This element is absolutely absent in the type m2 of Z. turicensis. As a result, in buccal view (Fig. 3B), in the unworn teeth, the interlophids are half stuffed up by enamel pillars (crescentoids and central conules), showing shallow interlophid notches, and in buccal and lingual views (Fig. 3B, C), the lophids display blunt peaks. Furthermore, the lophids 2 and 3 exhibit slight alternative positions and slight chevroned patterns, which is very uncommon in the gracile Z. turicensis and the other mammutid taxa. Similar morphology is also shown in Miomastodon tongxinensis (Fig. 3D, E). Therefore, a better solution is to restrict the genus name Zygolophodon only to the gracile morph-type, and attribute the robust type, e.g., S. gobiensis and Mio. tongxinensis, to another genus.
3.1.2 Comparison to Miomastodon merriami
Osborn (1921) reported Mastodon merriami, a pair of lower m3 and portions of two upper tusks from Virgin Valley Formation, Nevada, USA. Subsequently, Osborn (1922) established the new genus Miomastodon for his Mastodon merriami, that is, Mio. merriami (Osborn, 1921). Osborn (1936) attributed Mio. proavus from Pawnee Creek, Colorado, USA, published by Frick (1933), to Mio. merriami, and placed the hypodigm of Mastodon proavusCope, 1873 (a P4 and a fragment of an M1) to Serridentinus. However, later researchers often synonymized Mastodon merriami Osborn, 1921, and Mastodon proavusCope, 1873, with Z. proavus (Cope, 1873) (Madden and Storer, 1985; Lofgren and Anand, 2011). Although the specific name “proavus Cope, 1873” was published earlier than “merriami Osborn, 1921”, the characters of Mastodon proavus are obscure, and its hypodigm was possibly heterogeneous (Madden and Storer, 1985). The M1 of Mastodon proavus is to a large degree incomplete, and was presumed to belong to a bunodont taxon; and the P4, more possibly belonging to a zygodont taxon, possesses very little morphological information for defining a species. Here we regard Mastodon proavusCope, 1873, as nomen dubium.
The type m3 of Mastodon merriami Osborn, 1921 (Fig. 4A-C), although moderately to deeply worn, processes thicker enamel walls than that of typical Zygolophodon turicensis. The posttrite half lophids are also mesiodistally compressed, leaving mesiodistally wide interlophids, but the posttrite half lophids are relatively blunt. Although the posterior pretrite central conules seem to be absent, the anterior and posterior pretrite crescentoids are remarkably thicker than those of Z. turicensis. Therefore, like Serridentinus gobiensis and Miomastodon tongxinensis, the molar morphology of Mastodon merriami is more bunodont than that of Z. turicensis. As a result, the genus Miomastodon seems to be valid and should be revived for those untypical “Zygolophodon” with more or less bunodont molar morphology, such as Mio. merriami, Mio. gobiensis, and Mio. tongxinensis. Here, we regard each of these species as valid taxa (see the differential diagnosis).
3.1.3 Comparison to Gomphotherium productum and the other related Gomphotherium species
Miomastodon gobiensis was originally attributed to Serridentinus Osborn, 1923. Although Serridentinus has been synonymized with Gomphotherium, it is necessary to compare Miomastodon gobiensis with species that had previously been assigned into Serridentinus.
The type species of Serridentinus was Mastodon productusCope, 1875, which was established based on an incomplete mandible (AMNH 14383) from Santa Fé, New Mexico, USA. Osborn (1923) transferred this species to his newly established genus Serridentinus, i.e., S. productus (Cope, 1875), along with several North American species. Tobien (1972) synonymized Serridentinus with Gomphotherium, and attributed almost all of the North American species into one species G. productum. It should be mentioned that Tobien’s G. productum is a complex that contains more than one species. For simplicity, we compare Miomastodon gobiensis only with the type specimen of Mastodon productusCope, 1875. In G. productum (Fig. 4L-N), the posttrite main cuspid and mesoconelet are clearly separated, but they are not subdivided. The anterior and posterior crescentoids are short or absent, and the anterior and posterior central conules are strong and even subdivided as serration. The interlophids are relatively mesiodistally narrower than those of Mio. gobiensis. In buccal view (Fig. 4M), the interlophids are nearly fully stuffed up by enamel pillars rather than half stuffed up as in Mio. gobiensis. The interlophids are rather narrow. Generally speaking, the molar morphology of G. productum is rather bunodont, and that of Mio. gobiensis is intermediate between the bunodont G. productum and the zygodont Z. turicensis.
Osborn (1936) also attributed several Eurasian Trilophodon (= Gomphotherium) species into Serridentinus. The most important one is perhaps Mastodon angustidens forma subtapiroidesSchlesinger, 1917, from Eibiswald, Austria, which is nowadays referred to as Gomphotherium subtapiroideum (Schlesinger, 1917). G?hlich (2010) published abundant material of G. subtapiroideum from Sandelzhausen, and Wang et al. (2017) published a new species, G. tassyi, of which the tooth morphology is closely related to G. subtapiroideum.
In the lower molars of Gomphotherium subtapiroideum and G. tassyi (Fig. 4O-T), in occlusial view (Fig. 4O, R), the anterior and posterior crescentoids are present, with moderately developed (sometimes subdivided) posterior central conules. This feature is similar to that of Miomastodon gobiensis. The posttrite half lophids are moderately mesiodistally compressed, leaving relatively mesiodistally wide interlophids, also similar to those of Mio. gobiensis. However, the posttrite main cuspid and mesoconelet are less well subdivided than those of Mio. gobiensis. Alternative position and chevron structure are absent in G. subtapiroideum and G. tassyi, either. In buccal view (Fig. 4P, S), the interlophids are half stuffed up by the enamel pillars, as are those of Mio. gobiensis. Generally speaking, the molar of Mio. gobiensis resembles that of G. subtapiroideum and G. tassyi more than the other gomphotheriid and mammutid species.
3.1.4 Biometric comparison of molars
The m3s of the typical zygodont (Mammut and Zygolophodon, Fig. 5A) and bunodont taxa (Gomphotherium productum and G. angustidens, Fig. 5A) are clearly separated in the length-width plot. However, samples of Miomastodon and G. subtapiroideum/tassyi are scattered along the boundary between the ranges of zygodont and bunodont taxa (Fig. 5A) and overlap with each other. In the plot of W/L1+2-Wpo2/W2 (Fig. 5B), which enables the comparison of the m1-3 in the same panel, the sample of the zygodont taxa (Fig. 5B) is clustered at the top right of the panel and that of the G. angustidens (Fig. 5B) at the bottom left. However, the samples of Miomastodon, G. subtapiroideum/tassyi, and of G. productum (Fig. 5B) are located between the ranges of zygodont and G. angustidens samples, with slight overlaps at their boundary. Nevertheless, the samples of Miomastodon, G. subtapiroideum/tassyi, and G. productum cannot be separated from each other (Fig. 5B). The biometric comparison indicates that the width of the Miomastodon lower molar is intermediate between the typical zygodont and bunodont taxa, which is mainly contributed by the posttrite half lophid width. However, the biometric data of Miomastodon cannot be separated from G. subtapiroideum/tassyi and G. productum.
3.2 Comparison of the mandible of Miomastodon gobiensis to the other taxa
The mandible of Miomastodon merriami has been reported by Frick (1933) from Pawnee Creek, Colorado, and by Madden and Storer (1985) from Wood Mountain, Canada. A prominent difference between Mio. merriami and Mio. gobiensis is that the mandibular symphysis of Mio. merriami is shorter. The other features are very similar, including the slightly ventrally deflected mandibular symphysis, and the slightly dorsally bent mandibular tusk. Perhaps the most important feature shared with Mio. gobiensis is that the mandibular tusk is erected oval (see below).The mandible of Zygolophodon turicensis was rarely reported, except for a nearly complete mandible from Freising, Germany. However, the specimen had been ruined during World War II (G?hlich, 1998). Based on the descriptions and illustrations in the previous publications (see Osborn 1936:fig. 657; Lehmann, 1950:pl. 14, fig. 26), the mandibular tusk is laid oval cross-sectioned, unlike the erected oval cross-sectioned lower tusks in Miomastodon gobiensis. The laid oval cross-section of lower tusk is also known in Z. turicensis from Simorre, France (Tassy, 1977). The mandibular symphysis of the Freising Z. turicensis is moderately elongated as that of Mio. gobiensis (Fig. 5D). In lateral view, the mandibular symphysis of the Freising Z. turicensis is likewise slightly ventrally declined, but the ramus is lower than those of Mio. gobiensis.
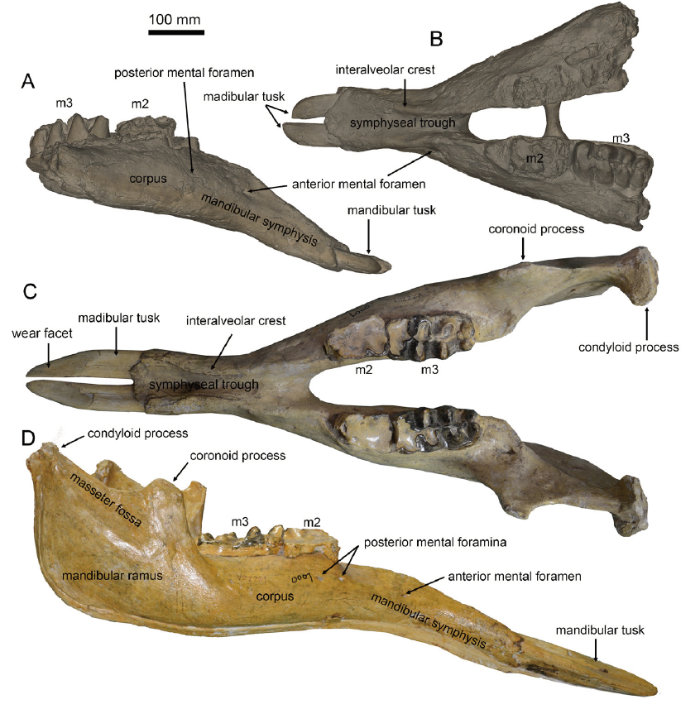
The mandible of Miomastodon gobiensis shared several features with Gomphotherium productum (Fig. 6A, B). Both possess long and rod-like mandibular tusks, and both have a moderately elongated mandibular symphysis. However, in G. productum, the cross-section of the mandibular tusk is also laid oval (Fig. 1G), as in Z. turicensis. In lateral view (Fig. 6A), the mandibular tusk of G. productum is more dorsally bent than that of Mio. gobiensis. In dorsal view (Fig. 6B), the mandibular symphysis of G. productum is slightly wider than that of Mio. gobiensis.
Fig. 6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Mandibles of Gomphotherium productum and G. tassyi, in comparison with Miomastodon gobiensis
A, B. AMNH 14383, the type mandible cast of G. productum, from Santa Fé, New Mexico, USA, in lateral (A) and dorsal (B) views; C, D. IVPP V 22781, unpublished mandible of G. tassyi, from the Heijiagou locality, Zhongning region, in dorsal (C) and lateral (D) views
The mandibular and tusk morphology of Gomphotherium subtapiroideum and G. tassyi is apparently distinct from those of Miomastodon gobiensis. In G. subtapiroideum and G. tassyi, the cross-section of mandibular tusk is pyriform with a clear dorsal groove. Furthermore, the mandibular symphysis of G. tassyi seems to be thicker and longer, and more ventrally inclined than that of Mio. gobiensis (Fig. 6C, D).
In the ancestral taxa of both the Gomphotheriidae and Mammutidae, for example, in the primitive mammutid Eozygodon, in the Gomphotherium annectens group, and in G. angustidens, the mandibular tusk is pyriform cross-sectioned with a strong or weak longitudinal dorsal groove, which is a plesiomorphy of elephantimorphs. This pyriform cross-sectioned mandibular tusk is also present in G. subtapiroideum and G. tassyi. In the derived taxa, for example, in the mammutid Miomastodon, Zygolophodon and gomphotheriid G. productum, the mandibular tusk is oval or circular. However, the cross-section of the lower tusk of Miomastodon differs from Zygolophodon and G. productum in having a more derived feature, the erected oval cross-section. This difference is clearly observed in the biometric plot (Fig. 5C), in which the samples of Mio. gobiensis and Mio. merriami species positions at the top left corner. This feature, the erected oval cross-sectioned lower tusk, is possibly an autapomorphy of the genus Miomastodon, which should be further attested in other Miomastodon species.
In biometric comparison of mandibular symphysis (Fig. 5D), Miomastodon gobiensis is positioned at the somewhat left of the bottom, indicating a very narrow and relatively short symphysis; while Mammut borsoni, being positioned at the leftmost and relatively lower part of the panel, has a shortest symphysis among all, and the symphysis is slightly wider than that of Miomastodon. The sample of Zygolophodon turicensis is closer to Mammut borsoni than to Miomastodon. Gomphotherium angustidens has the longest symphysis amongst them all, being at the right of the panel. Zygolophodon atavus and G. tassyi have mandibular symphysis that is intermediated between Miomastodon and G. angustidens. Whereas G. productum shows large variation in the biometric data, but generally, the symphysis of G. productum is relative wide.
3.3 Summary of morphological differences among Miomastodon gobiensis, Mio. merriami, Zygolophodon turicensis, Gomphotherium pruductum, and G. subtapiroideum
We integrate the key morphological points among Miomastodon gobiensis, Mio. merriami, Zygolophodon turicensis, Gomphotherium pruductum, and G. subtapiroideum/tassyi in Table 3. Based on the morphological comparison of the mandible and lower dentition, it can be inferred that the mandibular and lower dentition morphology of Mio. gobiensis and Mio. merriami is between Zygolophodon turicensis and several Gomphotherium species, e.g. G. productum and G. subtapiroideum/tassyi. For lower molar morphology, Mio. gobiensis and Mio. merriami resemble G. subtapiroideum/tassyi more than the other taxa, and they display intermediate morphology between the typical zygodont taxon Z. turicensis and the relatively bunodont taxon G. pruductum (the more typical bunodont taxa includes G. angustidens and G. annectens). In regards to mandibular morphology, Mio. gobiensis resembles that of Z. turicensis and the typical specimens of G. productum; however, seems more derived than that of G. subtapiroideum/tassyi. Furthermore, Mio. merriami possesses a shorter mandibular symphysis than Mio. gobiensis. One possible interpretation is that the length of the mandibular symphysis represents sexual dimorphism. This hypothesis cannot be verified, unless the two types of mandible are recovered from the same locality in the future. Another interpretation is that Mio. merriami and Mio. gobiensis represent distinct evolutionary directions. The former shortened their mandibular symphysis and tusks faster than the latter. Nevertheless, the similar tooth morphology strongly indicates that Serridentinus gobiensis could be attributed into Miomastodon. Furthermore, the presence of Miomastodon, as well as G. subtapiroideum/tassyi obscures the boundary of the Gomphotheriidae and Mammutidae. To further address this problem, it should be traced back to the early differentiation of the two families possibly during the Oligocene. As we know, the Oligocene elephantimorph Palaeomastodon also displays intermediate cheek tooth morphology between zygodonts and bunodonts, and was once considered an ancestor of the Mammutidae (Osborn, 1936). A collagen sequence phylogeny (Buckley et al., 2019) clusters Notiomastodon platensis (what we considered as gomphotheres) not with extant elephantids (believed to be derived from gomphotheres), but with Mammut americanum, which totally conflicts with the most common opinion of proboscidean evolution. It evokes the following speculation that gene introgression might occur between gomphotheres and mammutids.Table 3
Table 3Comparison of mandible and dentition among selected species of Zygolophodon, Miomastodon, and Gomphotherium
| Taxa | Zygolophodon turicensis | Miomastodon merriami | Miomastodon gobiensis | Gomphotherium productum complex | Gomphotherium subtapiroideum/tassyi |
|---|---|---|---|---|---|
| elongation of mandibular symphysis | moderately elongated | weakly elongated | moderately elongated | moderately elongated | moderately elongated |
| inclination of mandibular symphysis | slightly ventrally inclined | slightly ventrally inclined | slightly ventrally inclined | strongly ventrally inclined | strongly ventrally inclined |
| posterior mental foramina | ? | present | absent | present | present |
| cross-section of mandibular tusk | laid oval | erected oval | erected oval | laid oval to circular | pyriform |
| protrusion of mandibular tusk | short | short | short | short | long |
| enamel thickness | thin | thick | thick | thick | thick |
| mesiodistal distance of interlophids | wide | moderate | moderate | narrow | moderate |
| pretrite central conules | absent | absent or small | small and singular | large and serrate | small, singular or subdivided |
| posttrite half lophids | strongly subdivided | moderately subdivided | moderately subdivided | weakly subdivided or undivided | weakly subdivided or undivided |
| posttrite main cuspid and mesoconelet | separation clear or unclear | separation unclear | separation unclear | separation clear | separation clear |
| alternative position and chevron | absent | absent | present | absent | absent |
新窗口打开|下载CSV
3.4 Discussion of other related taxa
3.4.1 Serridentinus metachinjiensisOsborn, 1929, and Serridentinus chinjiensisOsborn, 1929Serridentinus metachinjiensisOsborn, 1929, was represented by a fragmentary right hemimandible with m2 and m3 (AMNH 19414) (Fig. 4D, E). The type locality is Chinji Bungalow, Pakistan, the Middle Miocene Chinji Formation. Tobien (1972) retained it within Gomphotherium, whereas Tassy (1983,1985) transferred it to Zygolophodon. The molar morphology of S. metachinjiensis is similar to that of Miomastodon gobiensis. In S. metachinjiensis (Fig. 4D, E), the pretrite crescentoids are thinner and central conules are smaller than those of Mio. gobiensis. However, the posttrite main cuspids and mesoconelets are slightly more bulbous and less subdivided than those of Mio. gobiensis, although these differences are minor. Serridentinus metachinjiensis might also represent a species that is similar to Gomphotherium subtapiroideum. However, the presence of slight alternative position and chevron of lophids leads us to revise it as Mio. metachinjiensis. Serridentinus chinjiensisOsborn, 1929 was represented by an incomplete right M3 (AMNH 19447) (Fig. 4I-K). It was also discovered from the Chinji Bungalow, the Chinji Formation, but the horizon seems slightly lower than that of S. metachinjiensis. Tassy (1983) synonymized S. chinjiensisOsborn, 1929, with Z. metachinjiensis (Osborn, 1929). Although fragmentary, the M3 exhibits typical zygodont features, such as the presence of clear pretrite crescentoids without central conules, highly subdivided posttrite half lophs with a sharp row of conelet summits (Fig. 4I), and the deep V-shaped interlophid notches in lateral view (Fig. 4J, K). Therefore, we would rather place S. chinjiensisOsborn, 1929, into Zygolophodon, and keep the validity of the specific name, Z. chinjiensis (Osborn, 1929).
3.4.2 Zygolophodon (?) junggarensis Chen, 1988
Chen (1988) erected this species based on fragmented upper jaws from the north of the Junggar Basin, China, Middle Miocene Halamagai Formation. Based on the plate and the description (Chen, 1988:pl.4, fig.3), it displays intermediate molar morphology between bunodonts and zygodonts. The posttrite lophs are moderately subdivided, the pretrite mesoconelets are bulbous, and pretrite crescentoids seem to be relatively thick. Therefore, this species should be attributed to Miomastodon. Further comparison is unavailable because few lower cheek teeth were known in Miomastodon. Here we temporary referred it to Miomastodon sp. However, the Junggar sample might be close to Mio. tongxinensis.
3.4.3 Mastodon atavusBorissiak, 1936
This species is represented by a nearly complete skeleton from the Early Miocene of Turgai, Kazakhstan. However, it lacks the braincase and upper jaws. Originally, Borissiak (1936) considered it to be closely related to Gomphotherium angustidens. Tassy (1985) and Tobien (1996) referred it to Zygolophodon atavus. The mandible is well preserved. The mandibular symphysis is slightly ventrally inclined and the mandibular tusks are rod-like with a ventrally inclined wear facet at the tip. However, the mandibular symphysis seems much longer than that of Z. turicensis (Borissiak, 1936:pl. 1, figs. 3, 4; also see Fig. 5D). The cross-section of the mandibular tusks is nearly circular, but with a slightly larger Dml than Ddv (laid oval; Fig. 6C; 55×52 mm). Unfortunately, the molars are fairly deeply worn, so that the crown morphology is not well-known (Borissiak, 1936:pl. 2, figs. 1, 2). The dimensions of the m3, and the width-length ratio of the first two lophids also falls into the ranges of Zygolophodon (Fig. 5A, B). Therefore, we retain this species to be Z. atavus, following Tassy (1985) and Tobien (1996).
4 Conclusions
In the present paper, Serridentinus gobiensisOsborn & Granger, 1932, Miomastodon tongxinensis Chen, 1978, and several related taxa are reevaluated, based on dental and mandibular features. We conclude that the genus Miomastodon Osborn, 1922, should be revived, including at least the following species: Mio. merriami (Osborn, 1921), Mio. tongxinensis Chen, 1978, Mio. gobiensis (Osborn & Granger, 1932), and Mio. metachinjiensis (Osborn, 1929). The mandibular tusk of Miomastodon is oval cross-sectioned, and the dorsoventral diameter is larger than the mediolateral diameter, which can be regarded as a generic autapomorphy (only known in Mio. merriami and Mio. gobiensis). Miomastodon, as a mammutid, possesses more bunodont cheek teeth than the contemporary Zygolophodon. However, the molar morphology of Miomastodon is similar to that of Gomphotherium subtapiroideum/tassyi, which possesses stronger mandibular symphysis and pyriform cross-sectioned lower tusk. The presence of Miomastodon and G. subtapiroideum/tassyi strongly suggests an involved evolutionary pattern existing between the Gomphotheriidae and Mammutidae.Acknowledgments
We thank Chen Guan-Fang, IVPP, China, for her extensive advice on this work. We thank J. Meng, AMNH, USA, P. Tassy, MNHN, France, U. G?hlich, NHMW, Austria, L. Costeur, NMB, Switzerland, G. R?ssner, SNSB-BSPG, Germany, R. Second and G. Corner, University of Nebraska State Museum of Natural History, USA, for allowing us access to the various taxa of proboscideans in the collection. We thank the two reviewers D. Mothé and Ji X P for their great advice on the original manuscript. We thank Y. O’Connor for the improvement of English. This work was supported by the Chinese Academy of Sciences (grant nos. XDB26000000, XDA20070203, QYZDY-SSW-DQC022, GJHZ1885), the National Natural Science Foundation of China (grant nos. 41872001, 41430102), and Special Research Program of Basic Science and Technology of the Ministry of Science and Technology (grant no. 2015FY310100).Supplementary material can be found at the website of Vertebrate PalAsiatica (http://english.ivpp.cas.cn/sp/PalAsiatica/vp_list/) in Vol. 58, Issue 2.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 5]
[本文引用: 1]
[本文引用: 10]
[本文引用: 4]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 6]
[本文引用: 4]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 9]
[本文引用: 6]
[本文引用: 2]
[本文引用: 12]
[本文引用: 8]
[本文引用: 19]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 6]
[本文引用: 6]
[本文引用: 4]
[本文引用: 4]
[本文引用: 1]
[本文引用: 3]
[本文引用: 11]
[本文引用: 2]
[本文引用: 3]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}